Do you want to buy antibiotics online without prescription? https://buyantibiotics24h.net/ - This is pharmacy online for you!
Pii: s0031-9384(01)00656-4
Physiology & Behavior 75 (2002) 237 – 241
Lingual and biting responses to selected lipids by the lizard
William E. Cooper Jr.a,*, Valentı´n Pe´rez-Melladob, Laurie J. Vittc
aDepartment of Biology, Indiana University-Purdue University at Fort Wayne, Fort Wayne, IN 46805, USA
bDepartamento de Biologia Animal, Universidad de Salamanca, Salamanca, Spain
cSam Noble Oklahoma Museum of Natural History and Department of Zoology, University of Oklahoma, 2401 Chautauqua Avenue,
Received 9 August 2001; received in revised form 25 September 2001; accepted 31 October 2001
Many lizards can identify food using chemical cues, but very little is known about the chemical constituents used for this purpose. We
experimentally investigated responses to several lipid stimuli by the omnivorous lacertid lizard Podarcis lilfordi, which had been shownpreviously to be capable of identifying prey using only chemical cues and to respond to pork fat by tongue-flicking and biting. In 60-s trialsin which stimuli were presented on cotton swabs, the lizards responded very strongly to pure pork fat and to oleic acid, but not to cholesterolor glycerol. Latency to bite swabs, the number of individuals that bit swabs, and the tongue-flick attack score, TFAS(R), which combineseffects of tongue-flicks and bites, showed stronger responses to fat than to cholesterol, glycerol, and distilled water but did not differsignificantly from responses to oleic acid. Several lines of evidence show that oleic acid elicited strong chemosensory and feeding responses. For individuals that did not bite, the number of tongue-flicks was significantly greater for oleic acid than for distilled water or glycerol, andnearly so for cholesterol. Latency to bite was significantly shorter for oleic acid than for distilled water, and TFAS(R) was significantlygreater for oleic acid than for distilled water and glycerol. In combination with pilot data indicating no strong response to the waxy, saturatedpalmitic acid, these findings suggest that oleic acid in particular and probably other unsaturated fatty acids found in animal fat contributestrongly to the food-related responses to lipids. D 2002 Elsevier Science Inc. All rights reserved.
Keywords: Behavior; Lipids; Food discriminations; Chemical senses; Squamata; Lacertidae; Podarcis lilfordi
tongue-flicking rates and permit identification of food asindicated by feeding attempts.
Squamate reptiles can identify food by tongue-flicking,
In the first study of responses to major categories of
but little is known about the types of chemicals that are
organic compounds found in foods by lizards, strongest
important in this regard beyond the finding that garter snakes
lingual and biting responses by the lacertid Gallotia caesaris
of the genus Thamnophis respond to specific proteins from
were detected for the carbohydrate sucrose and for fat, which
earthworms [1,2]. Among lizards, active foragers and omni-
contains a variety of lipids [9]. A second study revealed
vorous or herbivorous species can identify and evaluate foods
similar behavior by the lacertid Podarcis lilfordi, which
using chemical cues sampled by tongue-flicking (e.g., Refs.
additionally exhibited a stronger response to protein than to
[3 – 6]). In contrast to the extensive information now available
deionized water and a different sort of response to protein
about the relationships between foraging mode and prey
than to sucrose, which elicits licking [10]. The response to
chemical discrimination [3,7] and between plant diet and
protein was much weaker than to fat or sucrose. This contrasts
plant chemical discrimination [6,8], very little is known for
with findings for Thamnophis, in which specific proteins
lizards about the kinds of chemicals that stimulate increased
from earthworms, a favored food, elicit very strong responses[1,2]. The lack of strong response to protein by P. lilfordi maybe artifactual because the protein tested was bovine gammaglobulin rather than protein from the integument or exoskel-
* Corresponding author. Tel.: +1-219-481-6311; fax: +1-219-481-6087. E-mail address: cooperw@ipfw.edu (W.E. Cooper Jr.).
eton of typical prey. Additional studies are needed to ascer-
0031-9384/02/$ – see front matter D 2002 Elsevier Science Inc. All rights reserved. PII: S 0 0 3 1 - 9 3 8 4 ( 0 1 ) 0 0 6 5 6 - 4
W.E. Cooper Jr. et al. / Physiology & Behavior 75 (2002) 237–241
tain the types of lipids, carbohydrates, and proteins that may
pork fat was tested as a stimulus containing several types
contribute to chemical discriminations regarding prey and
of lipids. Oleic acid was used as an example of a non-
saturated fatty acid, glycerol as a component of triglycer-
Here, we report a first examination of the effects of a few
ides, and cholesterol as an additional lipid. After pilot tests
categories of lipids on chemosensory sampling and feeding
revealed no strong response to the waxy palmitic acid, this
behavior. Both G. caesaris and P. lilfordi responded very
saturated fatty acid was not included in the experiment.
strongly to pure pork fat, as indicated by biting attacks
Swabs were impregnated with stimuli as follows: Distilled
following chemical sampling by tongue-flicking [9,10].
water, glycerol, and oleic acid stimuli were prepared by
Because fat is composed primarily of trigylcerides, triglycer-
immersing the swab in the corresponding liquid; choles-
ides and their constituent molecules are the most likely
terol stimuli were prepared by immersing the swab in 5 ml
candidates as the effective compounds. The two major
of a solution of 0.025 g of cholesterol in glycerol. Pure
categories of fatty acids, saturated and unsaturated, seem
pork fat stimuli were prepared by inserting a swab into fat
likely to have different effects because at ambient temper-
at room temperature and wiping off excess with a paper
atures unsaturated fatty acids may be accessible as liquids,
towel. To human observers, the swabs prepared with all
whereas saturated fatty acids may be waxes. Glycerol, the
other major constituent of triglycerides, is potentially import-
Each trial was initiated by slowly moving a cotton swab
ant because it is ubiquitous. Other categories of lipids, such as
to a position 1.0 – 1.5 cm anterior to a lizard’s snout. Starting
sphingolipids, might also elicit strong responses whether or
with the first tongue-flick directed to the swab, the experi-
menter recorded the number of tongue-flicks directed to the
We experimentally studied responses to several lipids
swab in 60 s if the lizard did not bite. If the lizard bit, the
presented on cotton swabs to P. lilfordi, an actively foraging
latency to bite in seconds from the first tongue-flick until
omnivore. This species was used because it is known to be
the bite was recorded, as well as the number of tongue-
capable of prey chemical discrimination and to respond
strongly to fat [10,11] and is very abundant. Pork fat was
The experiment was conducted on 15 – 16 May 2001
used as a stimulus to estimate the strength of the full response
between 11:15 and 18:30 h. All data were collected by a
to a natural mixture of lipids, and water was used as an
single experimenter. Each lizard was tested with all stimuli
odorless control to determine response level in the absence of
in a repeated-measures (randomized blocks) design with a
any stimulus related to food. In addition, comparisons of
minimum intertrial interval of 30 min. To prevent possible
responses to a few readily available lipids (i.e., a single
bias that might occur if all lizards were tested using the
representative each of saturated and unsaturated fatty acids,
same sequence of stimuli, the stimulus sequence was varied
glycerol, and cholesterol) allowed us to determine whether
among individuals by incomplete counterbalancing in which
each type of lipid caused any increase in lingual and biting
one of the sequences starting with each stimulus type was
responses and the response strength relative to the full
Variables examined statistically were the number of
tongue-flicks, latency to bite, proportion of individuals thatbit, and TFAS(R), the tongue-flick attack score for repeated
measures [12]. The tongue-flick attack score gives the bestoverall indication of response strength by combining tongue-
Twenty adult male P. lilfordi were collected by noosing
flicks, which reflect chemosensory investigation, with biting,
and with traps baited with fruit on Aire, an islet offshore
which shows predatory attack [12 – 14]. In trials without bites,
from Menorca, Balearic Islands, Spain. The lizards were
TFAS(R) is the number of tongue-flicks. In trials with bites,
transported to a laboratory on Menorca, where they were
TFAS(R) is the maximum number of tongue-flicks in any trial
housed individually in transparent 40.5 Â 25.0 Â 26.5 cm
by the same individual plus 60 minus the latency to bite in
plastic terraria. Each terrarium contained a floor of indoor –
seconds. Thus, a bite is given heavier weight than any number
outdoor carpet and a water dish, and all sides were covered
with white paper to reduce disturbance to the lizards due to
We initially planned to analyze data on tongue-flicks,
movement by the experimenters in the room. The natural
latency to attack, and TFAS(R) using analysis of variance
photothermal cycle was supplemented by heat lamps, one at
for a single factor experiment having a randomized blocks
an end of each cage, which raised air temperature in cages to
design [15]. Because variances of all of these variables were
30 – 32 °C during testing and permitted thermoregulation.
significantly heterogeneous as indicated by Hartley’s Fmax
Lizards were left in cages overnight on the day of capture
tests, both for raw and logarithmically transformed data, we
and were tested on the following two days.
instead conducted nonparametric Friedman two-way ana-
Lipid stimuli were presented to lizards on the cotton tips
lyses of variance [16]. When significant main effects were
of 15-cm wooden applicators. Deionized water served as an
found, a procedure described in Ref. [16] for nonparametric
odorless control to assess response to the experimental
paired comparisons was used to test the significance of
milieu in the absence of stimuli relevant to food. Pure
differences between pairs of stimulus means.
W.E. Cooper Jr. et al. / Physiology & Behavior 75 (2002) 237–241
The main effect of stimulus condition on the proportion
of individuals that bit was assessed using a Cochran Q test
Biting responses by 18 P. lilfordi responding to chemical cues from fat(FAT), oleic acid (OLE), cholesterol (CHO), glycerol (GLY), and distilled
followed by sign tests of the differences between pairs of
water (WAT) on cotton swabs in 60-s trials
conditions [16]. Raw probabilities of the binomial tests are
reported, but a sequential Bonferroni procedure was used toevaluate significance [17]. All significance tests were two-
tailed, with a = .05. Two lizards failed to complete the
experiment, one ceasing responses to swabs after two trials,
the other after four. Thus, the sample size for all statistical
lizards that bit in neither the cholesterol nor the glycerol
condition, the number of tongue-flicks was greater tocholesterol for four individuals and for glycerol for the
Numbers of tongue-flicks were highly variable, espe-
other four ( P > 0.10). There was no difference between
cially in the oleic acid condition, which had the highest
numbers of tongue-flicks in the glycerol and water con-
mean values (Table 1). The lowest number of tongue-flicks
ditions when bites did not occur (nine individuals, choles-
occurred in the fat condition, but only because almost all
terol > water in six cases, water > cholesterol in three cases,
individuals bit too quickly to permit many tongue-flicks.
P > 0.10). Among 12 individuals that bit in neither the
There were no significant differences among conditions in
glycerol nor the distilled water condition, the number of
the number of tongue-flicks (c2 = 6.10, df = 4, P > .10).
tongue-flicks was greater in response to glycerol for seven
When the data are limited to trials in which bites did not
lizards, greater to water for four lizards, and there was one
occur, oleic acid elicited much greater numbers of tongue-
tie. This difference was not significant ( P > 0.10).
flicks than cholesterol, glycerol, and water (Table 1). Eight
Mean latency to bite varied greatly among conditions,
individuals did not bite in any of the oleic acid, glycerol,
being far shorter in the fat condition than in all others and
and water conditions, permitting sign tests. All eight indi-
somewhat shorter in the oleic acid condition than in the
viduals had higher numbers of tongue-flicks in the oleic acid
remaining conditions (Table 2). The main stimulus effect
condition than in either of the others. Numbers of tongue-
was highly significant (c2 = 30.55, df = 4, P < 1.0 Â 10 À 5).
flicks were significantly greater in response to oleic acid
Paired comparisons showed that latency to bite was signific-
than to glycerol or water (sign test, P < .0079 each) when no
antly shorter in response to fat than to cholesterol ( P < .05),
bite occurred. Using a Mann – Whitney U test to compare
glycerol ( P < .005), and distilled water ( P < .001). Despite
tongue-flicks between oleic acid and cholesterol when no
the substantially shorter mean latency to bite swabs bearing
bite occurred, there were six individuals that bit in the oleic
fat than oleic acid, this difference was not significant
acid condition, but not the cholesterol condition, and four
( P > 0.10). The only other significant difference between
individuals that bit in the cholesterol condition, but not in
pairs of conditions was the shorter latency to bite in the oleic
the oleic acid condition. For these individuals, the difference
acid condition than in the distilled water condition ( P < .05).
is not significant, but is marginal despite the small sample
Numbers of individuals that bit (Table 2) exhibited a
size (U = 5.0; n = 4, 6, P = .086), with greater tongue-flicks
similar pattern to latency to bite. The stimulus effect was
highly significant (c2 = 26.32, df = 4, P < .001). A signifi-
Similar comparisons among other pairs of stimuli in trials
cantly greater proportion of lizards bit in the fat condition
without bites were not significant, consistent with similarity
than in the cholesterol ( P =.002), glycerol ( P < .001), and
of their means (Table 1). In the comparison between eight
distilled water ( P < .00025). The only other substantial differ-ences were those between fat and oleic acid ( P < .016) andbetween oleic acid and water ( P < .032). The latter two
Table 1Mean tongue-flicks by P. lilfordi responding to chemical cues from fat
differences approached significance, but were not significant
(FAT), oleic acid (OLE), cholesterol (CHO), glycerol (GLY), and distilled
at the adjusted Bonferroni levels of .0072 and .0083, respect-
water (WAT) on cotton swabs in 60-s trials
ively, for the 10 possible comparisons.
TFAS(R) (Fig. 1) exhibited the same trends as the two
variables based solely on bites, but permitted slightly greater
resolution of differences among stimuli. The ranges of
TFAS(R) were 15 – 110 for fat, 14 – 97 for oleic acid, 2 – 110
for cholesterol, 2 – 77 for glycerol, and 1 – 64 for water.
Differences among stimuli were highly significant (c2 =
37.77, df = 4, P < 1 Â 10 À 5). Mean TFAS(R) was signific-
antly greater in response to fat than to cholesterol ( P < .05),
glycerol ( P < .005), and distilled water ( P < .001). TFAS(R)
W.E. Cooper Jr. et al. / Physiology & Behavior 75 (2002) 237–241
suggests that additional components of fat may contribute tothe overall response because even pure oleic acid, a super-normal stimulus probably never encountered naturally,appears to be slightly less effective than a natural lipidmixture. The ability of other triglycerides found in fat, suchas palmitoleic, linoleic, linolenic, and arachidonic acids, toinduce tongue-flicking and biting should be studied.
Cholesterol and glycerol had no significant effect on any
of the variables analyzed. The number of tongue-flicks forthese stimuli was very similar to that for distilled water. Although the difference was not significant, slightly moreindividuals bit and at shorter latency in response to choles-terol in glycerol than to distilled water, suggesting thatcholesterol or its combination with glycerol might have asmall effect requiring a larger sample size or a higher
Fig. 1. Mean tongue-flick attack scores for 18 adult male P. lilfordi in 60-s
concentration for its demonstration. Further hints of such
trials responding to cotton swabs bearing pure pork fat (FAT), oleic acid
an effect are that cholesterol in glycerol elicited a response
(OLE), cholesterol (CHO), glycerol (GLY), and distilled water (WAT).
2.23 times that to distilled water for TFAS(R) and that the
strength of response to the solution of cholesterol in glycerolwas slightly greater than half of that to fat, 0.55 for
was also significantly greater in the oleic acid condition than
TFAS(R). However, no effect was demonstrated, and the
in the glycerol ( P < .02) and distilled water conditions
lack of effect agrees with the finding that the cholesterol
( P < .001). No other differences were significant although
derivative estradiol-17b in peanut oil did not elicit a higher
those between cholesterol and distilled water and between
rate of tongue-flicking by male insectivorous lizards, Eume-
oleic acid and glycerol were substantial.
ces laticeps, than did peanut oil alone [18].
Tongue-flicking, which serves to sample chemicals for
analysis by vomerolfaction, and presumably stimulates gust-
atory responses as well, is a convenient empirically observ-able indicator of chemosensory investigation by lizards. It is
Lipids are a major stimulus contributing to prey chemical
also possible that the stimuli had different odors detected by
discriminations by the two species of lizards in which their
the lizards via olfaction, which can activate tongue-flicking
effectiveness has been examined and presumably in other
for vomerolfactory analysis [19]. Biting provides an indica-
lizards. As in previous studies of lacertid lizards [9,10], pure
tion of predatory attack that in most actively foraging,
pork fat elicited very strong responses from P. lilfordi.
insectivorous lizards occurs exclusively or nearly so as a
Although the number of tongue-flicks was slightly lower
response to prey chemicals (e.g., Refs. [20 – 22]). Such bites
in the fat condition than in the other conditions, this dif-
can readily be distinguished from defensive bites, which are
ference was not significant. When responding to fat stimuli,
not usually preceded by tongue-flicks in P. lilfordi, are
most of the lizards simply bit too quickly to permit many
accompanied by defensive postures, are brief, and are typ-
tongue-flicks. The high percentage of individuals that bit
ically followed by retreat of the lizard. Defensive tongue-
and the short latency to bite indicate that fat stimuli were
flicking may occur in some snakes, but is rare or absent in
lizards [23]. P. lilfordi is unusual among lizards in that it bites
Oleic acid elicited very strong lingual and biting respon-
more frequently in response to control stimuli such as water
ses. This is shown by the significantly greater TFAS(R) to
than do most lizards. This might be a consequence of intense
oleic acid than to glycerol and distilled water and the signifi-
competition for food in the very dense populations on Aire
cantly shorter latency to bite swabs bearing oleic acid than
[11] or of the omnivorous diet of this species.
distilled water. Although there were no significant differences
Tongue-flicks and bites are by far the most useful variables
among conditions in the number of tongue-flicks, the mean
for the study of prey chemical discriminations. In addition, a
number of tongue-flicks to oleic acid was more than twice that
shift in body position, especially elevation and/or turning of
for either glycerol or distilled water. For TFAS(R), response
the head, often accompanied by locomotory movement for a
strength to oleic acid was 2.95 times that to distilled water.
very short distance, sometimes occurs immediately following
These strong responses to oleic acid suggest that this abund-
tongue-flicks contacting a swab. Such behavior has been
ant constituent of mammalian fat may be an important
observed at low frequency in many lizard species (Cooper,
component of pork fat for its identification as a food.
unpublished). We observed such behavior in three individu-
Although the mean response strength to oleic acid was
als, one each in the oleic acid, cholesterol, and glycerol
statistically indistinguishable from that to pork fat, it was
conditions. The behaviors hint that detection of chemicals,
quantitatively only 0.73 of that to fat for TFAS(R). This
indicating the possible presence of food, may induce visual
W.E. Cooper Jr. et al. / Physiology & Behavior 75 (2002) 237–241
search for a prey item. In trials with presumptive search
[2] Wang D, Jiang XC, Chen P, Inouchi J, Halpern M. Chemical and
behavior, the greatest numbers of tongue-flicks in any trial
immunological analysis of prey-derived vomeronasal stimulants. Brain Behav Evol 1993;41:246 – 54.
were observed for glycerol (30) and cholesterol (39), and the
[3] Cooper WE. Foraging mode, prey chemical discrimination, and phy-
third greatest for oleic acid, indicating intense chemosensory
logeny in lizards. Anim Behav 1995;50:973 – 85.
investigation. For other individuals, prey chemicals on a
[4] Cooper WE. Correlated evolution of prey chemical discrimination
small white object, the swab, may suffice to release predatory
with foraging, lingual morphology, and vomeronasal chemoreceptor
abundance in lizards. Behav Ecol Sociobiol 1997;41:257 – 65.
[5] Cooper WE. Supplementation of phylogenetically correct data by two
The importance of lipids to chemosensory identification
species comparison: support for correlated evolution of foraging mode
of food by lizards seems clear, but the types of lipids that are
and prey chemical discrimination in lizards extended by first intra-
most important in this regard remain largely unknown. The
generic evidence. Oikos 1999;86:97 – 104.
present findings show that one specific unsaturated fatty
[6] Cooper WE. Convergent evolution of plant chemical discrimination by
acid elicits very strong responses, ones that could not be
omnivorous and herbivorous scleroglossan lizards. J Zool (London),in press.
distinguished statistically from that to a mixture of lipids in
[7] Cooper WE. An adaptive difference in the relationship between for-
fat. In addition, they hint that unsaturated fatty acids may be
aging mode and responses to prey chemicals in two congeneric scincid
among the primary effective compounds. However, the only
lizards. Ethology 2000;106:193 – 206.
saturated fatty acid tested was palmitic acid, which had no
[8] Cooper WE. Chemosensory discrimination of plant animal foods by
detectable effect on tongue-flicking and did not elicit bites
the omnivorous iguanian lizard Pogona vitticeps. Can J Zool 2000;78:1 – 5.
but is not a major constituent of animal fat [24].
[9] Cooper WE, Pe´rez-Mellado V. Chemosensory responses to sugar and
Adult P. lilfordi consume a wide variety of prey and plant
fat by the omnivorous lizard Gallotia caesaris with behavioral evidence
foods [11] that may contain a wide range of lipids. Oleic and
suggesting a role for gustation. Physiol Behav 2001;73:509 – 16.
other common fatty acids may be present in a wide range of
[10] Cooper WE, Pe´rez-Mellado V, Vitt LJ. Responses to major categories
food species. Similarities among them, especially among
of food chemicals by the lizard Podarcis lilfordi, in preparation.
[11] Pe´rez-Mellado V, Corti C. Dietary adaptations and herbivory in lacer-
unsaturated fatty acids, might permit identification of a wide
tid lizards of the genus Podarcis from western Mediterranean islands
range of food species, even foods previously unencountered.
(Reptilia: Sauria). Bonn Zool Beitr 1993;44:193 – 220.
Future studies should examine responses to additional animal
[12] Cooper WE, Burghardt GM. A comparative analysis of scoring meth-
fatty acids and to a range of lipids as yet untested, as well as to
ods for chemical discrimination of prey by squamate reptiles. J Chem
mixtures of lipid types. Although lipids may be adequate to
[13] Burghardt GM. Chemical-cue preferences of inexperienced snakes:
induce predatory attacks, their roles with respect to other
comparative aspects. Science 1967;157:718 – 21.
classes of organic chemicals remain unknown. Studies of the
[14] Burghardt GM. Chemical perception of reptiles. In: Johnston JW,
effects of combinations of proteins, lipids, and carbohydrates
Moulton DG, Turk A, editors. Communication by chemical signals.
found in preferred prey on lingually mediated chemosensory
New York: Appleton-Century-Crofts, 1970. pp. 241 – 308.
discriminations are needed to detect any synergisms or
[15] Winer BJ. Statistical principles in experimental design. New York:
[16] Zar JH. Biostatistical analysis. Upper Saddle River, NJ: Prentice-Hall,
[17] Wright SP. Adjusted p-values for simultaneous inference. Biometrics
[18] Cooper WE, Garstka WR, Vitt LJ. Female sex pheromone in the lizard
Eumeces laticeps. Herpetologica 1986;42:361 – 6.
This study was partially supported by project PB98-0270
[19] Cowles RB, Phelan RL. Olfaction in rattlesnakes. Copeia 1958;1958:
of the Spanish Ministry of Education and Culture, by the
Institut Menorqui d’Estudis, Consell Insular de Menorca,
[20] Cooper WE, Vitt LJ. Prey odor discrimination by the broad-headed
and by an international travel grant from Indiana University-
skink (Eumeces laticeps). J Exp Zool 1989;249:11 – 6.
Purdue University. Lizards were collected under a permit to
[21] Cooper WE, Hartdegen R. Lingual and biting responses to prey chem-
icals by ingestively naive scincid lizards: discrimination from control
VPM from the Servei de Biodiversitat, Conselleris d Medi
chemicals, time course, and effect of method of stimulus presentation.
[22] Cooper WE, Al-Johany AM, Vitt LJ, Habegger JJ. Responses to
chemical cues from animal and plant foods by actively foraging in-sectivorous and omnivorous scincine lizards. J Exp Zool 2000;287:
[23] Gove D. A comparative study of snake and lizard tongue-flicking,
[1] Schell PT, Powell R, Parmerlee JS. Notes on the natural history of
with an evolutionary hypothesis. Z Tierpsychol 1979;51:58076.
Ameiva chrysolaema (Sauria: Teiidae) from Barahona, Dominican
[24] Swenson MJ. Dukes’ physiology of domestic animals. 9th ed. Ithaca,
Republic. Copeia 1993;1993:859 – 62.
NY: Comstock Publishing Associates, 1977.
Sicherheitsdatenblatt gemäß 91/155/EWG Lt. § 14 Abs. (1) GefStoffV ist für Schädlingsbekämpfungsmittel kein Sicherheitsdatenblatt erforderlich. Ausstellungsdatum: 11.05.00 Ersatz für das Datenblatt vom: 07.09.99 n.a. = nicht anwendbar, n.v. = nicht vorhanden. Stoff-/Zubereitungs- und Firmenbezeichnung Angaben zum Produkt: Handelsname: Contrax-flüssig Angaben zum Hers
Wilderness and Remote First Aid Pre-Course Suggested Self-Study Material The American Red Cross Wilderness and Remote First Aid class is a hybrid course that builds on skills and knowledge offered in basic community level First Aid classes along with an introduction to some of the more advanced techniques presented in programs like BSA Lifeguard, American Red Cross Lifeguarding and Fir

 Physiology & Behavior 75 (2002) 237 – 241
Lingual and biting responses to selected lipids by the lizard
William E. Cooper Jr.a,*, Valentı´n Pe´rez-Melladob, Laurie J. Vittc
aDepartment of Biology, Indiana University-Purdue University at Fort Wayne, Fort Wayne, IN 46805, USA
bDepartamento de Biologia Animal, Universidad de Salamanca, Salamanca, Spain
cSam Noble Oklahoma Museum of Natural History and Department of Zoology, University of Oklahoma, 2401 Chautauqua Avenue,
Received 9 August 2001; received in revised form 25 September 2001; accepted 31 October 2001
Many lizards can identify food using chemical cues, but very little is known about the chemical constituents used for this purpose. We
experimentally investigated responses to several lipid stimuli by the omnivorous lacertid lizard Podarcis lilfordi, which had been shownpreviously to be capable of identifying prey using only chemical cues and to respond to pork fat by tongue-flicking and biting. In 60-s trialsin which stimuli were presented on cotton swabs, the lizards responded very strongly to pure pork fat and to oleic acid, but not to cholesterolor glycerol. Latency to bite swabs, the number of individuals that bit swabs, and the tongue-flick attack score, TFAS(R), which combineseffects of tongue-flicks and bites, showed stronger responses to fat than to cholesterol, glycerol, and distilled water but did not differsignificantly from responses to oleic acid. Several lines of evidence show that oleic acid elicited strong chemosensory and feeding responses.
Physiology & Behavior 75 (2002) 237 – 241
Lingual and biting responses to selected lipids by the lizard
William E. Cooper Jr.a,*, Valentı´n Pe´rez-Melladob, Laurie J. Vittc
aDepartment of Biology, Indiana University-Purdue University at Fort Wayne, Fort Wayne, IN 46805, USA
bDepartamento de Biologia Animal, Universidad de Salamanca, Salamanca, Spain
cSam Noble Oklahoma Museum of Natural History and Department of Zoology, University of Oklahoma, 2401 Chautauqua Avenue,
Received 9 August 2001; received in revised form 25 September 2001; accepted 31 October 2001
Many lizards can identify food using chemical cues, but very little is known about the chemical constituents used for this purpose. We
experimentally investigated responses to several lipid stimuli by the omnivorous lacertid lizard Podarcis lilfordi, which had been shownpreviously to be capable of identifying prey using only chemical cues and to respond to pork fat by tongue-flicking and biting. In 60-s trialsin which stimuli were presented on cotton swabs, the lizards responded very strongly to pure pork fat and to oleic acid, but not to cholesterolor glycerol. Latency to bite swabs, the number of individuals that bit swabs, and the tongue-flick attack score, TFAS(R), which combineseffects of tongue-flicks and bites, showed stronger responses to fat than to cholesterol, glycerol, and distilled water but did not differsignificantly from responses to oleic acid. Several lines of evidence show that oleic acid elicited strong chemosensory and feeding responses.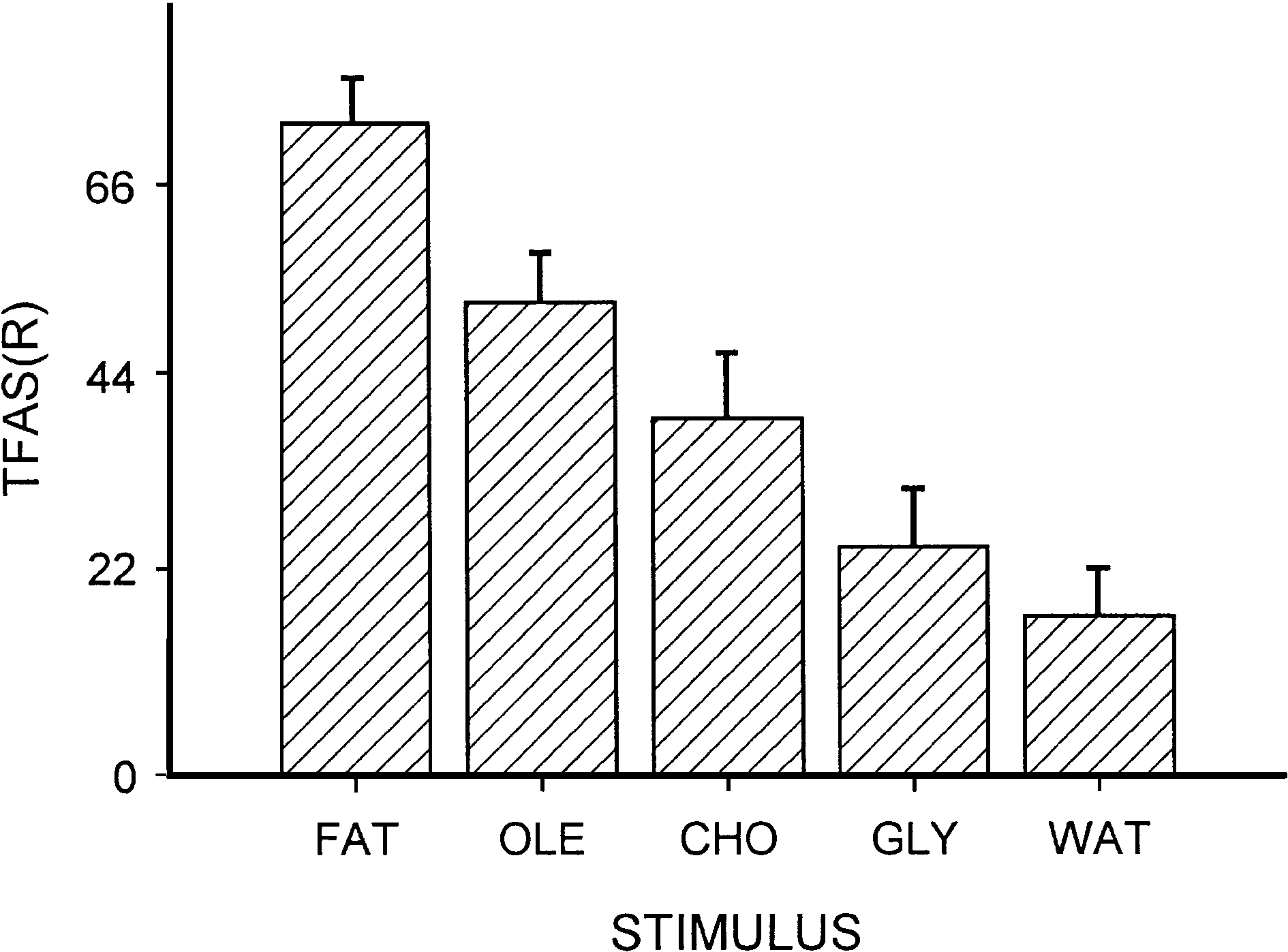 W.E. Cooper Jr. et al. / Physiology & Behavior 75 (2002) 237–241
suggests that additional components of fat may contribute tothe overall response because even pure oleic acid, a super-normal stimulus probably never encountered naturally,appears to be slightly less effective than a natural lipidmixture. The ability of other triglycerides found in fat, suchas palmitoleic, linoleic, linolenic, and arachidonic acids, toinduce tongue-flicking and biting should be studied.
W.E. Cooper Jr. et al. / Physiology & Behavior 75 (2002) 237–241
suggests that additional components of fat may contribute tothe overall response because even pure oleic acid, a super-normal stimulus probably never encountered naturally,appears to be slightly less effective than a natural lipidmixture. The ability of other triglycerides found in fat, suchas palmitoleic, linoleic, linolenic, and arachidonic acids, toinduce tongue-flicking and biting should be studied.