Do you want to buy antibiotics online without prescription? https://buyantibiotics24h.net/ - This is pharmacy online for you!
Functional interactions between dopamine, serotonin and norepinephrine neurons: an in-vivo electrophysiological study in rats with monoaminergic lesions
International Journal of Neuropsychopharmacology, Page 1 of 15. Copyright f 2008 CINP
Functional interactions between dopamine,
serotonin and norepinephrine neurons:an in-vivo electrophysiological studyin rats with monoaminergic lesions
Bruno P. Guiard1, Mostafa El Mansari1, Zul Merali1,2 and Pierre Blier1,2
1 Institute of Mental Health Research, University of Ottawa, Ottawa, Ontario, Canada2 Department of Cellular and Molecular Medicine, Faculty of Medicine ; Ottawa Health Research Institute, University of Ottawa,Ottawa, Ontario, Canada
Anatomical studies have established the existence of reciprocal relationships between the main populationof monoamine, serotonin (5-HT), norepinephrine (NE) and dopamine (DA) neurons in the brain. Thepresent study was thus conducted to examine the firing activity of 5-HT and NE neurons in DA-depletedrats, as well as the firing activity of DA neurons in 5-HT- or NE-depleted rats. The selective lesion of DAneurons elicited by 6-hydroxydopamine (6-OHDA) decreased the spontaneous firing activity of dorsalraphe (DR) nucleus 5-HT neurons by 60 %, thus revealing the excitatory effect of the DA input on these5-HT neurons. In contrast, the selective lesion of 5-HT neurons produced by 5,7-dihydroxytryptamine(5,7-DHT) enhanced by 36 % the firing activity of VTA DA neurons, thereby indicating an inhibitory effectof the 5-HT input on these DA neurons. With regard to the reciprocal interaction between DA and NEneurons, it was observed that the selective loss of DA neurons achieved by the intra-ventral tegmental area(VTA) injection of 6-OHDA increased the firing activity of a subset of locus coeruleus (LC) NE neurons by47 %. The selective loss of NE neurons in response to the intra-LC injection of 6-OHDA enhanced the firingactivity of VTA DA neurons by 70 %, demonstrating a net inhibitory role of the NE input on VTA DAneurons. These findings have important consequences for antidepressant treatments aimed at enhancingsimultaneously 5-HT, NE and DA transmission. Indeed, based on the understanding of such interactions,it may be possible to develop strategies to improve the effectiveness of antidepressant drugs by preventingcounter-productive negative feedback actions.
Received 12 August 2007 ; Reviewed 17 October 2007 ; Revised 21 November 2007 ; Accepted 5 December 2007
Key words : Antidepressants, dopamine, firing activity, norepinephrine, serotonin.
synthesis inhibition (Dremencov et al., 2007 ; Haddjeriet al., 1997 ; Reader et al., 1986). When NE neurons are
There are reciprocal projections between the major
lesioned, dorsal raphe (DR) 5-HT neurons discharge
groups of serotonin (5-hydroxytryptamine ; 5-HT) and
erratically at a low rate, but only for the first few days
norepinephrine (NE) neurons in the brain (Aston-
(Svensson et al., 1975). Although the loss of brain
Jones et al., 1991 ; Kaehler et al., 1999). The physio-
monoamine neurons does not necessarily reflect the
logical importance of such connections is evidenced by
pathophysiology of mood disorders, such an exper-
alterations in neuronal activity in lesion experiments.
imental approach can be initially used to establish the
When 5-HT neurons are lesioned, the firing rate of
net excitatory and/or inhibitory nature of a specific
locus coeruleus (LC) NE neurons is enhanced in a sus-
neurotransmitter at post-synaptic level. As an example
tained fashion by about 70 %, as is the case with 5-HT
of the clinical relevance of monoaminergic projections,selective 5-HT reuptake inhibition produces a marked
Address for correspondence : Dr B. P. Guiard, Ph.D., Institute of
inhibition of the spontaneous firing rate of LC NE
Mental Health Research (IMHR), 1145 Carling Avenue, University of
neurons (Dremencov et al., 2007 ; Seager et al., 2004,
Ottawa, Ottawa, K1Z 7K4, Ontario, Canada.
2005 ; Szabo et al., 2000). Low doses of atypical anti-
Tel. : +01 (613)-722-6521 (ext. 6732) Fax : +01 (613)-792-3935E-mail : bguiard@rohcg.on.ca
psychotics, which are now recognized as an effective
augmentation strategy in non-psychotic selective
and Esposito, 1995). However, electrical stimulation
serotonin reuptake inhibitor (SSRI)-resistant depressed
of the DR produces two different types of response
patients, reverse this inhibitory action via blockade of
in the VTA. Some DA neurons exhibit an inhibition-
5-HT2A receptors (Berman et al., 2007 ; Dremencov
excitation response while others show an initial
et al., 2007, Gharabawi et al., 2006a,b ; Rapaport et al.,
excitation followed by an inhibition (Gervais and
It is well documented that dopamine (DA) neurons
Descending pathways from the VTA also innervate
of the ventral tegmental area (VTA), giving rise to the
the LC (Ornstein et al., 1987). In-vivo recordings
mesolimbic/cortical DA system, send projections to
showed that direct iontophoretic application of DA in
the DR (Kale´n et al., 1988) and the LC (Beckstead et al.,
the LC of anaesthetized rats, suppresses the firing
1979), while in turn, receiving important inputs from
activity of NE neurons (Elam et al., 1986), while sys-
the latter nuclei (Herve´ et al., 1987). It therefore ap-
temic injection of the selective D2 antagonist halo-
pears crucial to examine the reciprocal interactions of
peridol enhances it (Piercey et al., 1994). In turn,
these three types of neurons to understand the effects
functional studies indicate that LC NE neurons
of medications acting on monoaminergic systems.
modulate DA neurons of the VTA. For instance, the
In particular, there is growing interest for DA in the
electrical stimulation of the LC as well as the systemic
field of mood disorders, since drugs that enhance its
administration of the selective NE reuptake inhibitor
transmission are clinically effective on their own. For
reboxetine, both increase NE levels in the VTA, pro-
example, the selective D2/D3 agonist pramipexole,
ducing excitation of DA neurons (Grenhoff et al., 1993 ;
customarily used in the treatment of Parkinson’s dis-
Linner et al., 2001). In contrast, the local application
ease (PD), was shown to be effective in depression
of NE in the VTA was shown to inhibit the electrical
as a monotherapy (Barone et al., 2006 ; Corrigan et al.,
activity of DA neurons (Aghajanian and Bunney, 1977 ;
2000), as well as an augmentation strategy for SSRI-
Grenhoff et al., 1995 ; White and Wang, 1984).
resistant patients (Goldberg et al., 2004 ; Lattanzi et al.,
In order to further elucidate the interactions be-
2002). Conversely, degeneration of DA neurons in PD
tween DA, 5-HT and NE neurons, the firing activity of
patients typically leads to anhedonia and loss of
5-HT and NE neurons was examined in DA-depleted
motivation, two symptoms frequently associated with
rats, as well as the firing activity of DA neurons in
depression (Harro and Oreland, 2001). More import-
antly, the prevalence of depression can reach 50 % inPD patients (McDonald et al., 2003). Taken together,
these observations suggest that an attenuation of DAtransmission could participate in the pathogenesis of
mood disorders, possibly in part through interactions
Male Sprague–Dawley rats (Charles River, St Constant,
with the 5-HT and/or the NE system(s).
QC, Canada) weighing 250–300 g, were used for the
There is consistent evidence regarding the dopa-
experiments. They were housed individually and kept
minergic regulation of DR 5-HT neurons. Infusion of
under standard laboratory conditions (12 : 12 h light/
the DA agonist apomorphine in the rat DR stimulates
dark cycle with free access to food and water). All
the firing rate of 5-HT neurons and the local release of
animals were handled according to the guidelines of
5-HT, while these effects are partially prevented by the
the Canadian Council on Animal Care (CCAC) and
selective D2 receptor antagonist raclopride (Ferre and
protocols in this study were approved by the local
Artigas, 1993 ; Martin-Ruiz et al., 2001). The hypothesis
Animal Care Committee (Ottawa Health Research
that DA interacts with 5-HT neurons, mainly through
activation of D2 receptors is also supported by the de-polarizing action of quinpirole, and its blockade with
the D2 receptor antagonist haloperidol, in rat 5-HTneurons recorded in vitro (Aman et al., 2006 ; Haj-
Rats were anaesthetized with a mixture 1 : 1 by volume
Dahmane, 2001). The exact nature of the effect of 5-HT
of xylazine (20 mg/ml) and ketamine (100 mg/ml)
on VTA DA neuron activity remains unclear, in that
and placed into a stereotaxic frame with atraumatic
both inhibitory and excitatory roles for 5-HT have
ear bars. To study interactions between 5-HT and DA
been observed. Acute intravenous administration of
neurons, rats were administered intracerebroven-
SSRIs, which probably enhances extracellular 5-HT
triculary (i.c.v., unilateral) with 5,7-dihydroxytrypt-
levels in the VTA, induces a small decrease in the firing
amine (5,7-DHT : 200 mg free base in 10 ml of 0.9 % NaCl
rate of VTA DA neurons (Di Mascio et al., 1998 ; Prisco
and 0.1 % ascorbic acid) or 6-hydroxydopamine
Interactions between monoaminergic neurons
(6-OHDA : 120 mg free base in 10 ml of 0.9 % NaCl and
0.1 % ascorbic acid) as previously described (Reader
The single-barrelled glass micropipettes were pos-
and Gauthier, 1984). The following stereotaxic co-
itioned using the following coordinates (in mm from
ordinates (in mm from bregma) : AP x0.9, L+1.5, V
lambda) : AP+1.0 to 1.2, L 0¡0.1, V 5–7. The presumed
3.7 were used to reach the lateral ventricle. The flow
5-HT neurons were then identified using the following
rate injection was 1 ml/min and after completion of the
criteria : a slow (0.5–2.5 Hz) and regular firing rate and
i.c.v. infusion of neurotoxins or vehicle, the syringe
long-duration (2–5 ms) bi- or triphasic extracellular
was left in place for 15 min to allow sufficient diffusion
waveform (Aghajanian and Vandermaelen, 1982).
before its withdrawal. One hour before the i.c.v. in-jection, animals lesioned with 5,7-DHT were pre-
treated with the selective NE reuptake inhibitordesipramine (25 mg/kg i.p.) and the selective DA
The single-barrelled glass micropipettes were pos-
reuptake inhibitor GBR12909 (25 mg/kg i.p.) to pre-
itioned using the following coordinates (in mm from
vent loss of NE and DA neurons, respectively. Those
Bregma) : AP x6 to x5.4, L 1 to 0.6, V 7–9. The pre-
lesioned with 6-OHDA were pre-treated with desipra-
sumed DA neurons were identified according to the
mine (25 mg/kg i.p.) and the SSRI fluoxetine (10 mg/
well-established electrophysiological properties in
kg i.p.) to prevent loss of NE and 5-HT neurons.
vivo : a typical triphasic action potential with a marked
Control rats (sham-operated) were subjected to the
negative deflection ; a characteristic long duration
same procedure and received the corresponding pre-
(>2.5 ms) often with an inflection or ‘notch ’ on the
treatments 1 h before the unilateral injection of 10 ml
rising phase ; a slow spontaneous firing rate (0.5–5 Hz)
with an irregular single spiking pattern with slow
To study interactions between central NE and DA
bursting activity (characterized by spike-amplitude
neurons, rats received a bilateral injection of 6-OHDA
decrement) (Grace and Bunney, 1983). As previously
(5 mg free base in 0.5 ml of 0.9 % NaCl and 0.1 % ascor-
described, a criterion of duration (>1.1 ms from the
bic acid) into the LC or VTA to limit the diffusion of
start of the action potential to the negative trough) was
the neurotoxin throughout the brain and consequently
produce a more selective deafferentation (Reader,
1982). This is of particular interest since intracerebralinjection of 6-OHDA may deplete both NE and DA
The single-barrelled glass micropipettes were pos-
levels (Reader and Gauthier, 1984). The following co-
itioned using the following coordinates (in mm from
ordinates were used : AP x1.1, L 1.1, V 5.5 for the LC
lambda) : AP x1.0 to x1.2, L 1.0–1.3, V 5–7. Spon-
(in mm from lambda) and AP – 5.8, L 0.7, V 8.5 for the
taneously active NE neurons were identified using the
VTA (in mm from bregma). Rats that received intra-
following criteria : regular firing rate (0.5–5.0 Hz) and
LC 6-OHDA were pre-treated, 1 h before, with fluoxe-
positive action potential of long duration (0.8–1.2 ms)
tine (10 mg/kg i.p.) and GBR12909 (25 mg/kg i.p.) and
exhibiting a brisk excitatory response to a nociceptive
those that received intra-VTA 6-OHDA, were ad-
pinch of the contralateral hind paw (Aghajanian and
ministered fluoxetine (10 mg/kg i.p.) and desipramine
Vandermaelen, 1982). The compression lasted y1 s
(25 mg/kg i.p.). It is noteworthy that intracerebral
with equal pressure being applied to the paw of rats ;
administration of 6-OHDA was reported to be more
once the opposite sides of the forceps made contact
effective in depleting NE than the systemic treatment
with each other, the forceps were then released. Of
with DSP4 (Lookingland et al., 1986).
interest, it has also been reported that the numberof elicited bursts is largely independent of paw-compression intensity.
Ten days after the injection of the neurotoxins, ratswere anaesthetized with chloral hydrate (400 mg/kg
The firing patterns of DA and NE neurons (both
i.p.) and placed into a stereotaxic frame. The extra-
displaying a bursting activity) were analysed by spike-
cellular recordings of the 5-HT, DA and NE neurons in
interval burst analysis following the criteria estab-
the DR, VTA and LC, were performed using single-
lished by Grace and Bunney (1984). The onset of a
barrelled glass micropipettes (R&D Scientific Glass,
burst was defined as the occurrence of two spikes
Spencerville, MD, USA) preloaded with a 2 M NaCl
with an inter-spike interval shorter than 0.08 s. The
solution. Their impedance typically ranged between
termination of bursts was defined as an inter-spike
interval (ISI) of o0.16 s. The detailed analysis of ISI for
DA, NE and also 5-HT neurons in sham-operated and
lesioned rats is provided in Supplementary material
Neurochemical analyses of the neurotoxic lesions
Rats treated with the i.c.v. injection of 6-OHDA dis-
Biochemical analysis of brain monoamine levels
played a 70 % reduction of DA levels in the striatum
The effectiveness and selectivity of the neurochemical
(Table 1a). In the DA neuron-lesioned rats, no changes
lesions was confirmed by measuring 5-HT, NE and DA
in 5-HT levels were detected in the frontal cortex
concentrations at specific brain sites. The frontal cortex
(Table 1a), the hippocampus and the striatum (data
and striatum (including the nucleus accumbens) were
not shown) compared to sham-operated rats. More-
chosen to determine the extent of 5-HT and DA de-
over, no changes in NE levels were reported in these
pletion, respectively, as preferential serotonergic and
post-synaptic structures (Table 1a, and data not
dopaminergic projections from the DR and the VTA.
shown) with the exception of the frontal cortex
The hippocampus was selected to examine NE con-
(0.13¡0.02 ng/mg vs. 0.22¡0.01 ng/mg in lesioned
centration since it has been repeatedly shown to have
and sham-operated rats, respectively ; p<0.05), sug-
high levels of this monoamine (Dailly et al., 2006).
gesting that desipramine does not effectively protect
Immediately after electrophysiological experiments
NE terminals within the latter region. The lesion of
sham-operated rats and lesioned rats were sacrified,
5-HT neurons induced by the i.c.v. injection of 5,7-
the brain removed and stored at x80 xC. The frontal
DHT significantly reduced the levels of 5-HT in the
cortex, hippocampus and striatum were dissected as
frontal cortex by 87 %. The selectivity of the 5,7-DHT
previously described (Chenu et al., 2006). Each separ-
lesion was confirmed from the observations that the
ate brain area was placed in an Eppendorf tube with
concentrations of NE and DA were not different be-
tween lesioned and sham-operated rats in the frontal
an internal standard, dihydroxybenzylamine), homo-
cortex, hippocampus and striatum (Table 1a, and data
genized using ultrasound and centrifuged at 8000 g for
15 min. The supernatant was analysed for monoamine
The lesion of VTA DA neurons elicited by local
content using high-performance liquid chromato-
injection of 6-OHDA produced a significant reduction
in DA levels (48 %) in the striatum (Table 1b). In-terestingly, the depletion of VTA DA neurons pro-
duced a similar degree of depletion (50 %) in the frontalcortex of lesioned rats compared to sham-operated
Electrophysiological data were expressed as mean¡
rats (0.08¡0.02 ng/mg vs. 0.19¡0.03 ng/mg ; p<0.01).
S.E.M of the firing rate, number of single spikes, number
5-HT and NE levels were unchanged in the frontal
of bursts and single spikes per burst. Statistical com-
cortex, hippocampus and striatum of rats that received
parisons among DR, VTA and LC of sham-operated
the intra-VTA injection of 6-OHDA with respect to the
and lesioned rats were performed using two-tailed
sham-operated animals (Table 1b, and data not
Student’s t tests. The means (number¡S.E.M) of neurons
shown). The lesion of LC NE neurons achieved by
recorded per track in sham-operated and lesioned rats
local injection of 6-OHDA significantly decreased the
were also compared using a two-tailed Student’s t test.
level of NE in the hippocampus by 66 % (Table 1b).
For the lesioning studies, each neurotransmitter peak
5-HT and DA levels were not different between sham-
from the HPLC was converted into values represent-
operated and lesioned rats in the frontal cortex, hippo-
ing ng/mg wet weight tissue based on external
campus and striatum (Table 1b, and data not shown).
neurotransmitter standards of that day. A Student’st test was used to analyse between-group differences.
Effect of DA neuron lesion on the firing activity of 5-HTneurons in the DR
Desipramine hydrochloride, GBR12909 and the neuro-toxins (5,7-DHT creatinine sulphate, 6-OHDA hydro-
The mean number of DR 5-HT neurons recorded per
bromide) were purchased from Sigma-Aldrich (St
track was not significantly different between sham-
Louis, MO, USA). Fluoxetine was purchased from
operated (n=24 tracks) and DA neuron-lesioned rats
Medisca Pharmaceutic Inc. (Montreal, Canada) and
(n=22 tracks, Table 2). The mean firing frequency
idazoxan from Sigma/RBI (Oakville, ON, Canada). All
of DR 5-HT neurons in DA neuron-lesioned rats was
neurotoxins were dissolved before experiments and
significantly decreased by 60 % compared to sham-
protected from light during the injection.
Interactions between monoaminergic neurons
Table 1. Effect of monoaminergic neurons lesion on the extent of 5-HT, NE and DA levels in the frontal cortex,hippocampus and striatum
(a) Intracerebroventricular injections of neurotoxinsSham-operated rats
(b) In-situ injections of neurotoxinsSham-operated rats
VTA, Ventral tegmental area ; LC, locus coeruleus. Values are expressed as the mean¡S.E.M. (ng/mg wet tissue) of each monoamine in sham-operated rats and lesioned rats, 10 dafter the injection of vehicle or the neurotoxin. One hour before the administration of the neurotoxin, fluoxetine, desipramineand/or GBR12909 were administered to protect the 5-HT, NE and DA, respectively and enhance the selectivity of the lesion. * p<0.05 and ** p<0.01 relative the corresponding group of sham-operated rats.
Table 2. Electrophysiological characteristics of monoaminergic neurons in sham-operated and lesioned rats
DR, Dorsal raphe ; VTA, ventral tegmental area ; LC, locus coeruleus. * p<0.05, ** p<0.01 and *** p<0.001 relative the corresponding group of sham-operated rats.
Effect of 5-HT neuron lesion on the firing activity of DA
sham-operated (n=49 tracks) and 5-HT neuron-
lesioned rats (n=61 tracks, Table 2). In rats with their5-HT neurons lesioned, the mean firing frequency of
The mean number of VTA DA neurons recorded
VTA DA neurons was significantly increased by 36 %
per track was not significantly different between
compared to sham-operated rats (Figure 2c). In order
Figure 1. Effect of dopaminergic lesion with6-hydroxydopamine (6-OHDA) on the electrophysiologic
Figure 2. Effect of serotonergic lesion with 5,7-
activity of 5-HT neurons in the dorsal raphe (DR).
dihydroxytryptamine (5,7-DHT) on the electrophysiological
Examples of typical recordings of DR 5-HT neurons obtained
activity of DA neurons in the ventral tegmental area (VTA).
in (a) a sham-operated rat and (b) a DA neuron-lesioned
Examples of typical recordings of VTA DA neurons obtained
rat. (c) Mean¡S.E.M. of frequency (Hz) of 5-HT neurons
in (a) a sham-operated rat and (b) a 5-HT neuron-lesioned rat,
recorded in the DR of sham-operated rats (n=5)
each asterisk indicates a burst. (c) Mean¡S.E.M. of frequency
(grey histogram) and 6-OHDA-treated rats (n=6)
(Hz) of DA neurons. Number of single spikes/min (d),
(white histogram). *** p<0.001 indicates significantly
bursts/min (e), and single spikes/burst (f), recorded in the
different from sham-operated rats. The numbers within the
VTA of sham-operated rats (n=7) (grey histogram) and
histograms indicate the number of neurons recorded.
5,7-DHT-treated rats (n=7) (white histogram). * p<0.05,** p<0.01 and *** p<0.001 indicates significantly differentfrom sham-operated rats. The numbers within the histograms
to determine whether this increase was due to
indicate the number of neurons recorded.
an alteration of single spike and/or burst activity, amore detailed analysis was performed. The meannumber of single spikes/min did not quite reach
tracks) than in VTA-lesioned rats (n=36 tracks,
the pre-determined level of statistical significance
Table 2). The mean firing frequency of all spontaneous
in depleted rats compared to sham-operated rats
active LC NE neurons was significantly increased by
(Figure 2d). However, the number of bursts/min and
47 % in VTA-lesioned rats compared to sham-operated
of single spikes per burst was significantly increased
in rats with 5-HT neurons lesioned (Figure 2e, f).
(Dremencov et al., 2007), it was found that 77 % of NEcells discharged only in single-spike mode while the
Effect of DA neuron lesion on the firing activity of NE
rest displayed bursting activity. These percentages
were not affected by the lesion of VTA DA neurons
The mean number of NE neurons recorded per track
since 71 % and 29 % of LC NE neurons exhibited a
was significantly higher in sham-operated rats (n=10
single spike and bursting activity, respectively. The
Interactions between monoaminergic neurons
Table 3. Effect of VTA DA neuron lesion on the sensory-evoked firing activity of LC NE neurons
VTA, Ventral tegmental area ; LC, locus coeruleus. * p<0.05 and ** p<0.01, relative the sham-operated group of rats.
mean firing frequency of NE cells discharging with a
significant difference between depleted rats and sham-
single spike pattern, was significantly increased by
operated rats was detected in the mean number of
33 % in VTA DA neuron-lesioned rats compared to
single spikes/min (Figure 5d). However, the number
sham-operated rats (Figure 3c). In addition, among the
of bursts/min and single spikes per burst were sig-
LC NE cells displaying a bursting activity, the mean
nificantly increased in LC NE neuron-lesioned rats
firing frequency of LC NE neurons was increased by
compared to sham-operated rats (Figure 5e, f ).
59 % in VTA DA neuron-lesioned rats compared tosham-operated (Figure 4c). No significant differencewas detected between both groups of rats in the mean
number of single spikes/min ; while the number of
The present electrophysiological study showed that
bursts/min and single spikes per burst were signifi-
lesioning 5-HT and NE neurons increased the firing
cantly increased in VTA-lesioned rats (Figure 4e, f ).
activity of VTA DA neurons. Conversely, lesioning
In addition, it has been observed that the percentage
DA neurons decreased DR 5-HT neuronal firing but
of neurons displaying a sensory-evoked burst firing is
significantly increased in VTA DA neuron-lesionedrats compared to sham-operated rats, while no differ-ences were detected in the number of spikes per burst
Reciprocal interactions between DA and 5-HT
In VTA-lesioned rats, pre-treatment with the selec-
The lesion of DA neurons by i.c.v. administration of
tive a2-adrenoreceptor antagonist idazoxan (1 mg/kg
6-OHDA produced a profound and selective decrease
i.v.) did not increase the mean number of NE neurons
in brain DA levels (Table 1). In these experimental
recorded per track compared with the corresponding
conditions, the discharge rate of DR 5-HT neurons
group of VTA DA neuron-lesioned rats receiving no
was reduced by 60 % indicating that DA input exerts
pre-treatment (0.7¡0.1, n=12 tracks vs. 0.6¡0.1, n=61
a tonic excitatory effect on 5-HT neurons in intact brain.
tracks, respectively). However, the mean firing fre-
It may seem unusual that despite the marked attenu-
quency of LC NE neurons after the idazoxan pre-
ation of DR 5-HT neuronal firing in lesioned rats, the
treatment was significantly increased (4.5¡0.3 Hz,
number of DR 5-HT neurons recorded per track was
n=9 neurons vs. 3.1¡0.3 Hz, n=24 neurons ; p=
unchanged. However, previous electrophysiological
data indicate that conditions that decrease the firingactivity of DR 5-HT neurons by about 50 % do not
Effect of NE neuron lesion with 6-OHDA on the firing
necessarily modify the number of neurons found per
track (Blier et al., 1986). These findings are consistent
The mean number of DA neurons recorded per track
with previous in-vivo electrophysiological and neuro-
was not significantly different between sham-operated
chemical studies having shown that the systemic ad-
(n=49 tracks) and LC NE neuron-lesioned rats (n=64
ministration of the non-selective DA receptor agonist
tracks, Table 2). In the lesioned rats, the mean firing
apomorphine, increases the firing rate of 5-HT neurons
frequency of VTA DA neurons was increased by 70 %
(Martin-Ruiz et al., 2001), thereby enhancing 5-HT
compared to sham-operated rats (Figure 5c). No
outflow in the rat DR (Ferre et al., 1994 ; Ferre and
Figure 3. Effect of dopaminergic lesion with
6-hydroxydopamine (6-OHDA) on the electrophysiological
activity of locus coeruleus (LC) NE neurons discharging in asingle-spike mode. Examples of typical recordings of LC NE
Figure 4. Effect of dopaminergic lesion with
neurons obtained in (a) a sham-operated rat and (b) a ventral
6-hydroxydopamine (6-OHDA) on the electrophysiological
tegmental area (VTA) DA neuron-lesioned rat. The
activity of locus coeruleus (LC) NE neurons exhibiting a
characteristic increase in firing activity (without producing a
single-spike and burst activity. Examples of typical
burst) followed by a ‘ silence’ in response to pinch is
recordings of LC NE neurons obtained in (a) a sham-operated
indicated by an arrow. (c) Mean¡S.E.M. of frequency (Hz) of
rat and (b) a ventral tegmental area (VTA) DA neuron-
NE neurons recorded in the VTA of sham-operated rats
lesioned rat, each asterisk indicates a burst. The characteristic
(n=3) (grey histogram) and 6-OHDA-treated rats (n=7)
increase in firing activity followed by a ‘silence’ in response
(white histogram). * p<0.05 indicates significantly different
to pinch is indicated by an arrow. (c) Mean¡S.E.M. of
from sham-operated rats. The numbers within the histograms
frequency (Hz) of NE neurons. Number of single spikes/
indicate the number of neurons recorded.
min(d), bursts/min (e), and single spikes/burst (f), recordedin the VTA of sham-operated rats (n=3) (grey histogram)and 6-OHDA-treated rats (n=7) (white histogram). * p<0.05,** p<0.01 and *** p<0.001 indicates significantly different
Artigas, 1993 ; Martin-Ruiz et al., 2001). Likewise, it has
from sham-operated rats. The numbers within the histograms
been shown by intracellular recordings that both ap-
indicate the number of neurons recorded.
plication of DA and the D2/D3 agonist quinpirole in DRslices produces a concentration-dependent membranedepolarization of 5-HT neurons (Haj-Dahmane, 2001). These D2-like receptors are probably located on the
antagonists 6,7-dinitroquinoxaline-2,3-dione and 2-
5-HT neurons themeselves, because the effects of quin-
amino-5-phosphonopentanoic acid, did not prevent
pirole were unaffected by tetrodotoxin (which blocks
the DA-induced depolarization of DR 5-HT membrane
neuronal conduction ; Haj-Dahmane, 2001). Further-
(Haj-Dahmane, 2001) ruling out the possibility that
more, the pharmacological inactivation of ionotropic
the excitatory effects of DA involved a local release
and metabotropic glutamate receptors by the selective
of glutamate. It is well known that the DR is driven by a
Interactions between monoaminergic neurons
results indicate that the DA input exerts a direct excit-
atory effect, probably via D2 receptors on DR 5-HT
neurons while the influence of other non-dopaminergic
The i.c.v. injection of 5,7-DHT produced a robust
and selective decrease in brain 5-HT levels (Table 1)
leading to a 36 % increase in the discharge rate of VTA
DA neurons. This enhancement of DA neuronal ac-
tivity resulted from a higher number of bursts and
spikes per burst. The putative inhibitory effect of the
5-HT input on DA neurons suggested by the present
study is consistent with the finding that SSRIs, which
probably raise extracellular 5-HT levels in the VTA,
induce a slight decrease in the firing rate of VTA DAneurons (Di Mascio et al., 1998 ; Prisco and Esposito,
1995). The inhibitory influence of the 5-HT input on
DA neurons was further supported by the observation
that low doses of the 5-HT1A receptor agonist
8-OHDPAT, known to attenuate the electrical activity
of DR 5-HT neurons, increases the firing rate and the
number of burst discharge of DA neurons in the VTA
(Arborelius et al., 1993 ; Lejeune and Millan, 1998,
2000 ; Lejeune et al., 1997), and consequently DA
release at the somatodendritic (Chen and Reith, 1995)
and terminal levels (Ago et al., 2002 ; Arborelius et al.,
1993 ; Rasmusson et al., 1994 ; Tanda et al., 1994). How-
Figure 5. Effect of noradrenergic lesion with 6-
ever, high doses of 8-OHDPAT which preferentially
hydroxydopamine (6-OHDA) on the electrophysiological
activate post-synpatic receptors, also produce in-
activity of DA neurons in the ventral tegmental area (VTA).
creases in the discharge of DA neurons (Lejeune and
Examples of typical recordings of VTA DA neurons obtained
Millan, 1998). Consequently, the inhibitory effect of the
in (a) a sham-operated rat and (b) a locus coeruleus (LC)
5-HT input on VTA DA neurons remains debatable.
NE neuron-lesioned rat, each asterisk indicating a burst.
Indeed, initial intracellular recordings showed that 5-
(c) Mean¡S.E.M. of frequency (Hz) of DA neurons. Number
HT depolarized 46 % of rat VTA DA neurons (Pessia
of single spikes/min (d), bursts/min (e), and single
et al., 1994) and stimulated the release of [3H]DA in
spikes/burst (f), recorded in the VTA of sham-operated rats
VTA slices (Beart and McDonald, 1982). In line with
(n=6) (grey histogram) and 6-OHDA-treated rats (n=7)
these results, microinfusion of 5-HT in the VTA in-
(white histogram). ** p<0.01 and *** p<0.001 indicates
creased DA release in projection areas such as the nu-
significantly different from sham-operated rats. The numberswithin the histograms indicate the number of neurons
cleus accumbens (Guan and McBride, 1989). It was
also observed that the electrical stimulation of the DRproduces two different types of response in the VTA :some cells exhibit an inhibition-excitation response
noradrenergic input ; therefore a lesion of DA neurons
while other DA neurons show an excitation followed
might alter 5-HT neuronal activity indirectly via its in-
by an inhibition (Gervais and Rouillard, 2000). These
teractions with NE neurons. The possibility that the
results raised the possibility that the regulation of VTA
i.c.v. injection of 6-OHDA destroyed NE terminals in
DA neurons involves various post-synaptic 5-HT re-
the DR should be considered since a reduction of NE
ceptors. The 5-HT2A and 5-HT2C receptor subtypes
levels has been detected in the hippocampus (Table 1,
identified in the VTA (Cornea-Hebert et al., 1999 ;
n.s.) and the frontal cortex (data not shown, p<0.05) in
Nocjar et al., 2002) are of particular interest since their
lesioned rats. However, Svensson et al. (1975) demon-
pharmacological activation respectively stimulates and
strated that the spontaneous firing rate of DR 5-HT
suppresses, VTA DA neuronal activity (Di Giovanni
neurons after 6-OHDA is unaltered suggesting that
et al., 2000 ; Di Matteo et al., 2000 ; Gobert et al., 2000 ;
a putative loss of NE would not have a sustained im-
Millan et al., 2000 ; Pessia et al., 1994 ; Prisco et al.,
pact on 5-HT neuronal activity. In conclusion, these
1994) and DA release in the nucleus accumbens
(De Deurwaerdere et al., 2004 ; Porras et al., 2002).
neuronal activity (Nilsson et al., 2005 ; Piercey et al.,
Overall, it would thus seem that the inhibitory influ-
1994). An involvement of 5-HT neurons in the disin-
ence of 5-HT input plays a predominant role in
hibitory effect of DA neuron lesion on NE neuronal
the regulation of DA neuronal activity given that the
firing should also be considered. Indeed, the decrease
systemic administration of the non-selective 5-HT2
in DR 5-HT neuronal activity in response to the VTA
receptor antagonist ritanserin dose-dependently in-
lesion could lead to a significant reduction of 5-HT
creases the burst firing and the firing rate of VTA DA
release in the LC, thus contributing to the increase of
neurons (Ugedo et al., 1989). Importantly, the lesion of
NE neuronal firing through a lower tonic activation
5-HT neurons might have disrupted more than the
of 5-HT2A receptors (Szabo and Blier, 2002). Such an
serotonergic inputs to the VTA. Growing evidence
indirect mechanism, as well as a removal of the direct
suggests that feedback loops involving the 5-HT sys-
inhibitory effect of DA in the LC, could explain the
tem may control VTA DA neuronal activity. For ex-
present findings that the VTA lesion increased the
ample, it was proposed that the activation of 5-HT1A
firing activity of some NE neurons. Despite the elev-
and 5-HT2A receptor subtypes in the medial prefrontal
ated discharge of LC NE neurons observed in VTA-
cortex produces an excitation of VTA DA neurons
lesioned rats, the number of NE neurons recorded
(Bortolozzi et al., 2005 ; Diaz-Mataix et al., 2005, 2006).
was significantly reduced. Since, a partial loss of NE
However, in contrast to the present study, a lower to-
neurons was unlikely on the basis of the neuro-
nic stimulation of these pathways in 5-HT-depleted
chemical analysis (Table 1), the possibility that NE
rats should have produced an attenuation of the VTA
neurons were tonically inhibited by a local increase of
DA neurons’ firing activity. It may also be claimed that
NE levels was considered. In VTA-lesioned rats, the
the excitatory effect of the DR 5-HT lesion on VTA DA
systemic administration of the a2-adrenoreceptor an-
neurons was indirectly mediated by NE neurons. In-
tagonist idazoxan did not allow the recording of
deed, it is well established that NE neurons send pro-
more NE neurons. It thus seems possible that DA
jections to the VTA and that a lesion of 5-HT neurons
could also impinge on LC afferents, such as glutama-
increases the firing activity of the LC NE neurons
tergic neurons (Nilsson et al., 2005) that could in-
(Haddjeri et al., 1997). This is, however, unlikely since
directly participate in activating, at least in part, a
the present study suggests that NE exerts a robust in-
subpopulation of LC NE neurons. In line with this
hibitory action on VTA DA neurons (Figure 5).
hypothesis, it was reported that the stimulation ofthe VTA can produce an activation of NE neuronalactivity (Deutch et al., 1986). It should also be men-
Reciprocal interactions between DA and NE neurons
tioned that the intra-VTA injection of 6-OHDA could
The lesion of VTA DA neurons obtained with local
have destroyed, at least partly, NE terminals in the
injection of 6-OHDA resulted in a significant and
VTA. Although, this possibility cannot be completely
selective reduction of striatal DA levels (Table 1). This
excluded, it is noteworthy that no reductions of the
selective lesion also results in a 47 % increase in the
NE levels were detected either in the hippocampus
discharge rate of LC NE neurons suggesting that
(Table 1) or in the frontal cortex or striatum of lesioned
DA exerts an inhibitory action on these NE neurons.
rats (data not shown). Consequently, the present find-
Although the majority of LC NE neurons discharge
ings suggest that DA exerts a direct inhibitory action
spontaneously in a single-spike mode (Dawe et al.,
on some LC NE neurons through D2 receptors as well
2001 ; Dremencov et al., 2007), about 30 % of NE
as an indirect activation involving other neuronal
neurons exhibit both single and burst patterns of
spontaneous firing. In the later population of NE
The lesion of LC induced by the local injection of
neurons, VTA lesion also elevated the mean discharge
6-OHDA resulted in a significant and selective re-
rate as the result of a significant increase in the number
duction of brain NE levels (Table 1). This lesion in-
of bursts and spikes per burst. The putative inhibitory
creased the discharge rate of VTA DA neurons by 70 %
influence of DA input upon LC NE neurons is in line
owing to a significant higher number of bursts and
with the observation that direct iontophoretic appli-
action potentials per burst. This finding was consistent
cation of DA suppresses the firing rate of NE neurons
with those of earlier studies having shown that
(Elam et al., 1986). Similarly, in-vivo extracellular re-
the systemic administration of low doses of the a2-
cordings have demonstrated an involvement of D
adrenoreceptor agonist clonidine, which attenuates
ceptors in this inhibitory effect, since the systemic
overall NE transmission (Haddjeri and Blier, 2000 ;
administration of antipsychotic drugs, including the
Haddjeri et al., 1998 ; Szabo and Blier, 2001), also in-
creases the firing activity of VTA DA neurons
2 receptor antagonist haloperidol, increased LC NE
Interactions between monoaminergic neurons
(Georges and Aston-Jones, 2003 ; Gobbi et al., 2001 ;
VTA DA neurons, it seems highly unlikely that the
Millan et al., 2000). Taken together, these data strongly
decrease in burst firing activity in LC NE-lesioned rats
suggest that NE inputs exert an inhibitory influence
altered those pathways. However, it is more conceiv-
on spontaneous VTA DA neuronal activity. In line
able that the depletion of NE reduces the electrical
with this assumption, initial electrophysiological
activity of LDT GABAergic neurons projecting to the
studies have demonstrated that the microiontophoretic
VTA. These hypotheses will be of interest to address
application of NE in the VTA reduces the firing of
in future investigations. Nevertheless, so far a direct
DA neurons while this effect is blocked by the non-
attenuation of post-synaptic D2 receptor stimulation
selective a1/2-adrenoreceptor antagonist piperoxane
seems a more plausible hypothesis to explain the
(White and Wang, 1984). Thus far, it has been shown
increase in DA neuronal activity observed in LC-
that the stimulation of a1-adrenoceptors exerts a direct
excitatory influence on VTA DA neurons and an
The present lesion experiments emphasize the com-
indirect inhibitory effect by activating GABA inter-
plex regulation of 5-HT, NE and DA neuronal firing
neurons (Grenhoff et al., 1995 ; Steffensen et al., 1998).
activity. Further electrophysiological studies in com-
Thus, an attenuation of GABA release in the VTA,
bination with local or systemic administration of
could support the observation reported herein that DA
pharmacological agents will be useful in clarifying the
neuronal activity is enhanced in LC-lesioned rats.
pharmacological bases for these interactions in vivo.
With regard to a2-adrenoceptors, it has been demon-
Indeed, a better knowledge of such interactions could
strated that the local application of clonidine in the
provide important information for improving the
VTA does not inhibit DA neurons (Aghajanian and
treatment of depression, more specifically for pharma-
Bunney, 1977) ruling out the possibility that the
cotherapies aimed at enhancing simultaneously DA,
inhibitory effects of NE involved post-synaptic a2-
NE and 5-HT transmission without triggering counter-
adrenoceptors. However, divergent results have also
productive negative feedback actions. It is noteworthy
been reported. For example, the systemic adminis-
that the most of the traditional antidepressant drugs
tration of idazoxan or selective NE reuptake inhibitors,
tend to increase the endogenous monoaminergic
which raise extracellular NE levels in the VTA has
tone whereas the present study is based on the effect
been shown to increase the burst firing activity of DA
of the removal of this tone. However, those lesioning
neurons in the VTA (Grenhoff and Svensson, 1989,
experiments may initiate novel treatment approaches
1993 ; Linner et al., 2001 ; Shi et al., 2000). Clearly, fur-
whose clinical utility remains to be demonstrated.
ther studies are needed to determine the complexmechanism by which the NE input regulates VTA DA
neuronal activity. In particular, it would be relevantto address the issue of the selectivity of NE since the
Supplementary material accompanies this paper on
inhibitory effect of NE was found to be prevented
the Journal’s website (http://journals.cambridge.org).
by the iontophoretic application of the D2 receptorantagonist sulpiride (White and Wang, 1984). Indirect
mechanisms might also be involved in the effect ofLC NE neuron depletion on VTA DA neuronal
This study was supported by the Canadian Institutes
activity. The laterodorsal tegmentum (LTD), an ad-
for Health Research grant (G6152147) and salary sup-
jacent region to the LC that projects heavily to the
port from the University of Ottawa Institute of
VTA (Forster and Blaha, 2000 ; Oakman et al., 1995 ;
Mental Health Research to B.G, M.E., Z.M. and P.B.,
Omelchenko and Sesack, 2005) is mainly concerned.
as well as a Research Chair in Psychopharmacology
Indeed, inputs to the VTA that arrive from the LTD
from the Canadian Government to P.B.
components which have the ability to regulate not
only DA neuron population activity but also their
burst firing (Lodge and Grace, 2006, Maskos et al.,2005). Interestingly, evidence indicates that NE excitescholinergic and non-cholinergic neurons in the LTD
(Kohlmeier and Reiner, 1999 ; Koyama and Sakai, 2000)
Adell A, Artigas F (2004). The somatodendritic release
suggesting that an attenuation of NE transmission in
of dopamine in the ventral tegmental area and its
the LTD may affect the activity of VTA. As cholinergic
regulation by afferent transmitter systems.
and glutamatergic neurons exert excitatory effects on
Neuroscience & Biobehavioral Reviews 28, 415–431.
Aghajanian GK, Bunney BS (1977). Dopamine’
area of rats treated systemically with (+/x)-8-hydroxy-2-
autoreceptors’ : pharmacological characterization by
(di-n-propylamino)tetralin. Journal of Neurochemistry 64,
microiontophoretic single cell recording studies.
Naunyn Schmiedebergs Archives of Pharmacology 297, 1–7.
Chenu F, Dailly E, Bourin M (2006). Effect of antidepressant
Aghajanian GK, Vandermaelen CP (1982). Intracellular
drugs on 6-OHDA-treated mice in the FST. European
identification of central noradrenergic and serotonergic
Neuropsychopharmacology 17, 187–193.
neurons by a new double labeling procedure. Journal of
Cornea-Hebert V, Riad M, Wu C, Singh SK, Descarries L
(1999). Cellular and subcellular distribution of the
Ago Y, Sakaue M, Baba A, Matsuda T (2002).
serotonin 5-HT2A receptor in the central nervous system
Selective reduction by isolation rearing of 5-HT1A
of adult rat. Journal of Comparative Neurology 409, 187–209.
receptor-mediated dopamine release in vivo in
Corrigan MH, Denahan AQ, Wright CE, Ragual RJ, Evans
the frontal cortex of mice. Journal of Neurochemistry 83,
DL (2000). Comparison of pramipexole, fluoxetine, and
placebo in patients with major depression. Depression &
Aman TK, Shen RY, Haj-Dahmane S (2006). D2-like
dopamine receptors depolarize dorsal raphe serotonin
Dailly E, Chenu F, Petit-Demouliere B, Bourin M (2006).
neurons through the activation of nonselective cationic
Specificity and efficacy of noradrenaline, serotonin
conductance. Journal of Pharmacology and Experimental
depletion in discrete brain areas of Swiss mice by
neurotoxins. Journal of Neuroscience Methods 150, 111–115.
Arborelius L, Chergui K, Murase S, Nomikos GG, Hook BB,
Dawe GS, Huff KD, Vandergriff JL, Sharp T, O’Neill MJ,
Chouvet G, Hacksell U, Svensson TH (1993). The
Rasmussen K (2001). Olanzapine activates the rat locus
5-HT1A receptor selective ligands, (R)-8-OH-DPAT and
coeruleus : in vivo electrophysiology and c-Fos
(S)-UH-301, differentially affect the activity of midbrain
immunoreactivity. Biological Psychiatry 50, 510–520.
dopamine neurons. Naunyn Schmiedeberg’s Archives of
De Deurwaerdere P, Navailles S, Berg KA, Clarke WP,
Spampinato U (2004). Constitutive activity of the
Aston-Jones G, Shipley MT, Chouvet G, Ennis M, van
serotonin2C receptor inhibits in vivo dopamine release
Bockstaele E, Pieribone V, Shiekhattar R, Akaoka H,
in the rat striatum and nucleus accumbens. Journal of
Drolet G, Astier B, et al. (1991). Afferent regulation of locus
coeruleus neurons : anatomy, physiology and
Deutch AY, Goldstein M, Roth RH (1986). Activation
pharmacology. Progress in Brain Research 88, 47–75.
of the locus coeruleus induced by selective stimulation
Barone P, Scarzella L, Marconi R, Antonini A, Morgante L,
of the ventral tegmental area. Brain Research 363, 307–314.
Bracco F, Zappia M, Musch B (2006). Pramipexole versus
Di Giovanni G, Di Matteo V, Di Mascio M, Esposito E
sertraline in the treatment of depression in Parkinson’s
(2000). Preferential modulation of mesolimbic vs.
disease : a national multicenter parallel-group randomized
nigrostriatal dopaminergic function by serotonin(2C/2B)
study. Journal of Neurology 253, 601–607.
receptor agonists : a combined in vivo electrophysiological
Beart PM, McDonald D (1982). 5-Hydroxytryptamine and
and microdialysis study. Synapse 35, 53–61.
5-hydroxytryptaminergic-dopaminergic interactions in
Di Mascio M, Di Giovanni G, Di Matteo V, Prisco S,
the ventral tegmental area of rat brain. Journal of Pharmacy
Esposito E (1998). Selective serotonin reuptake inhibitors
reduce the spontaneous activity of dopaminergic neurons
Beckstead RM, Domesick VB, Nauta WJ (1979). Efferent
in the ventral tegmental area. Brain Research Bulletin 46,
connections of the substantia nigra and ventral tegmental
area in the rat. Brain Research 175, 191–217.
Di Matteo V, Di Giovanni G, Di Mascio M, Esposito E
Berman RM, Marcus RN, Swanink R, McQuade RD,
(2000). Biochemical and electrophysiological evidence
Carson WH, Corey-Lisle PK, Khan A (2007). The efficacy
that RO 60-0175 inhibits mesolimbic dopaminergic
and safety of aripiprazole as adjunctive therapy in
function through serotonin(2C) receptors. Brain Research
major depressive disorder : a multicenter, randomized,
double-blind, placebo-controlled study. Journal of Clinical
Diaz-Mataix L, Artigas F, Celada P (2006). Activation of
pyramidal cells in rat medial prefrontal cortex projecting
Blier P, de Montigny C, Azzaro AJ (1986). Effect of
to ventral tegmental area by a 5-HT1A receptor agonist.
repeated amiflamine administration on serotonergic and
European Neuropsychopharmacology 16, 288–296.
noradrenergic neurotransmission : electrophysiological
Diaz-Mataix L, Scorza MC, Bortolozzi A, Toth M, Celada P,
studies in the rat CNS. Naunyn Schmiedeberg’s Archives of
Artigas F (2005). Involvement of 5-HT1A receptors in
prefrontal cortex in the modulation of dopaminergic
Bortolozzi A, Diaz-Mataix L, Scorza MC, Celada P, Artigas F
activity : role in atypical antipsychotic action. Journal of
(2005). The activation of 5-HT receptors in prefrontal cortex
enhances dopaminergic activity. Journal of Neurochemistry
Dremencov E, El Mansari M, Blier P (2007). Noradrenergic
augmentation of escitalopram response by risperidone :
Chen NH, Reith ME (1995). Monoamine interactions
electrophysiologic studies in the rat brain. Biological
measured by microdialysis in the ventral tegmental
Interactions between monoaminergic neurons
Elam M, Clark D, Svensson TH (1986). Electrophysiological
Grenhoff J, Nisell M, Ferre S, Aston-Jones G, Svensson TH
effects of the enantiomers of 3-PPP on neurons in the locus
(1993). Noradrenergic modulation of midbrain dopamine
coeruleus of the rat. Neuropharmacology 25, 1003–1008.
cell firing elicited by stimulation of the locus coeruleus in
Ferre S, Artigas F (1993). Dopamine D2 receptor-mediated
the rat. Journal of Neural Transmission (General Section) 93,
regulation of serotonin extracellular concentration in the
dorsal raphe nucleus of freely moving rats. Journal of
Grenhoff J, North RA, Johnson SW (1995). Alpha
1-adrenergic effects on dopamine neurons recorded
Ferre S, Cortes R, Artigas F (1994). Dopaminergic regulation
intracellularly in the rat midbrain slice. European Journal of
of the serotonergic raphe-striatal pathway : microdialysis
studies in freely moving rats. Journal of Neuroscience 14,
Grenhoff J, Svensson TH (1989). Clonidine modulates
dopamine cell firing in rat ventral tegmental area. European
Forster GL, Blaha CD (2000). Laterodorsal tegmental
Journal of Pharmacology 165, 11–18.
stimulation elicits dopamine efflux in the rat nucleus
Grenhoff J, Svensson TH (1993). Prazosin modulates the
accumbens by activation of acetylcholine and glutamate
firing pattern of dopamine neurons in rat ventral
receptors in the ventral tegmental area. European Journal of
tegmental area. European Journal of Pharmacology 233, 79–84.
Guan XM, McBride WJ (1989). Serotonin microinfusion into
Georges F, Aston-Jones G (2003). Prolonged activation of
the ventral tegmental area increases accumbens dopamine
mesolimbic dopaminergic neurons by morphine
release. Brain Research Bulletin 23, 541–547.
withdrawal following clonidine : participation of
Haddjeri N, Blier P (2000). Effect of neurokinin-I receptor
imidazoline and norepinephrine receptors.
antagonists on the function of 5-HT and noradrenaline
Neuropsychopharmacology 28, 1140–1149.
neurons. Neuroreport 11, 1323–1327.
Gervais J, Rouillard C (2000). Dorsal raphe stimulation
Haddjeri N, Blier P, de Montigny C (1998). Acute and
differentially modulates dopaminergic neurons in the
long-term actions of the antidepressant drug mirtazapine
ventral tegmental area and substantia nigra. Synapse 35,
on central 5-HT neurotransmission. Journal of Affective
Gharabawi G, Canuso C, Pandina G, Bossie C, Kujawa M,
Haddjeri N, de Montigny C, Blier P (1997). Modulation of
Kosik-Gonzalez M, Turkoz I, Mahmoud R, Shelton R
the firing activity of noradrenergic neurones in the rat
(2006a). Risperidone treatment of resistant depression :
locus coeruleus by the 5-hydroxtryptamine system.
a double-blind placebo randomized trial. Neuropsycho-
British Journal of Pharmacology 120, 865–875.
Haj-Dahmane S (2001). D2-like dopamine receptor activation
Gharabawi G, Canuso C, Pandina G, Bossie C, Kujawa M,
excites rat dorsal raphe 5-HT neurons in vitro. European
Kosik-Gonzalez M, Turkoz I (2006b). A double-blind
Journal of Neuroscience 14, 125–134.
placebo-controlled study of adjunctive risperidone for
Harro J, Oreland L (2001). Depression as a spreading
treatment-resistant major depressive disorder. European
adjustment disorder of monoaminergic neurons : a case for
primary implication of the locus coeruleus. Brain Research.
Gobbi G, Muntoni AL, Gessa GL, Diana M (2001).
Brain Research Reviews 38, 79–128.
Clonidine fails to modify dopaminergic neuronal activity
Herve´ D, Pickel VM, Joh TH, Beaudet A (1987). Serotonin
during morphine withdrawal. Psychopharmacology (Berlin)
axon terminals in the ventral tegmental area of the
rat : fine structure and synaptic input to dopaminergic
Gobert A, Rivet JM, Lejeune F, Newman-Tancredi A,
neurons. Brain Research 435, 71–83.
Adhumeau-Auclair A, Nicolas JP, Cistarelli L, Melon C,
Kaehler ST, Singewald N, Philippu A (1999). Dependence of
Millan MJ (2000). Serotonin(2C) receptors tonically
serotonin release in the locus coeruleus on dorsal raphe
suppress the activity of mesocortical dopaminergic and
neuronal activity. Naunyn Schmiedeberg’s Archives of
adrenergic, but not serotonergic, pathways : a combined
dialysis and electrophysiological analysis in the rat.
Kale´n P, Skagerberg G, Lindvall O (1988). Projections from
the ventral tegmental area and mesencephalic raphe to the
Goldberg JF, Burdick KE, Endick CJ (2004). Preliminary
dorsal raphe nucleus in the rat. Evidence for a minor
randomized, double-blind, placebo-controlled trial of
dopaminergic component. Experimental Brain Research 73,
pramipexole added to mood stabilizers for treatment-
resistant bipolar depression. American Journal of Psychiatry
Kohlmeier KA, Reiner PB (1999). Noradrenaline excites
non-cholinergic laterodorsal tegmental neurons via two
Grace AA, Bunney BS (1983). Intracellular and extracellular
distinct mechanisms. Neuroscience 93, 619–630.
electrophysiology of nigral dopaminergic neurons – 1.
Koyama Y, Sakai K (2000). Modulation of presumed
Identification and characterization. Neuroscience 10,
cholinergic mesopontine tegmental neurons by
acetylcholine and monoamines applied iontophoretically
Grace AA, Bunney BS (1984). The control of firing pattern in
in unanesthetized cats. Neuroscience 96, 723–733.
nigral dopamine neurons : burst firing. Journal of
Lattanzi L, Dell’Osso L, Cassano P, Pini S, Rucci P,
Houck PR, Gemignani A, Battistini G, Bassi A, Abelli M,
Cassano GB (2002). Pramipexole in treatment-resistant
coeruleus neurons by clozapine and haloperidol :
depression : a 16-week naturalistic study. Bipolar Disorder 4,
involvement of glutamatergic mechanisms. International
Journal of Neuropsychopharmacology 8, 329–339.
Lejeune F, Millan MJ (1998). Induction of burst firing
Nocjar C, Roth BL, Pehek EA (2002). Localization of
in ventral tegmental area dopaminergic neurons
5-HT(2A) receptors on dopamine cells in subnuclei of
by activation of serotonin (5-HT)1A receptors :
the midbrain A10 cell group. Neuroscience 111, 163–176.
WAY 100,635-reversible actions of the highly
Oakman SA, Faris PL, Kerr PE, Cozzari C, Hartman BK
selective ligands, flesinoxan and S 15535. Synapse 30,
(1995). Distribution of pontomesencephalic cholinergic
neurons projecting to substantia nigra differs significantly
Lejeune F, Millan MJ (2000). Pindolol excites dopaminergic
from those projecting to ventral tegmental area. Journal of
and adrenergic neurons, and inhibits serotonergic neurons,
by activation of 5-HT1A receptors. European Journal of
Omelchenko N, Sesack SR (2005). Laterodorsal tegmental
projections to identified cell populations in the rat ventral
Lejeune F, Newman-Tancredi A, Audinot V, Millan MJ
tegmental area. Journal of Comparative Neurology 483,
(1997). Interactions of (+)- and (x)-8- and 7-hydroxy-2-(di-
n-propylamino)tetralin at human (h)D3, hD2 and
Ornstein K, Milon H, McRae-Degueurce A, Alvarez C,
h serotonin1A receptors and their modulation of the
Berger B, Wurzner HP (1987). Biochemical and
activity of serotoninergic and dopaminergic neurones in
radioautographic evidence for dopaminergic afferents
rats. Journal of Pharmacology and Experimental Therapeutics
of the locus coeruleus originating in the ventral
tegmental area. Journal of Neural Transmission 70, 183–191.
Linner L, Endersz H, Ohman D, Bengtsson F, Schalling M,
Pessia M, Jiang ZG, North RA, Johnson SW (1994). Actions
Svensson TH (2001). Reboxetine modulates the firing
of 5-hydroxytryptamine on ventral tegmental area neurons
pattern of dopamine cells in the ventral tegmental area and
of the rat in vitro. Brain Research 654, 324–330.
selectively increases dopamine availability in the
Piercey MF, Smith MW, Lum-Ragan JT (1994). Excitation of
prefrontal cortex. Journal of Pharmacology and Experimental
noradrenergic cell firing by 5-hydroxytryptamine1A
agonists correlates with dopamine antagonist properties.
Lodge DJ, Grace AA (2006). The laterodorsal tegmentum is
Journal of Pharmacology and Experimental Therapeutics 268,
essential for burst firing of ventral tegmental area
dopamine neurons. Proceedings of the National Academy of
Porras G, Di Matteo V, Fracasso C, Lucas G, De
Deurwaerdere P, Caccia S, Esposito E, Spampinato U
Lookingland KJ, Chapin DS, McKay DW, Moore KE (1986).
(2002). 5-HT2A and 5-HT2C/2B receptor subtypes
Comparative effects of the neurotoxins N-chloroethyl-
modulate dopamine release induced in vivo by
N-ethyl-2-bromobenzylamine hydrochloride (DSP4) and
amphetamine and morphine in both the rat nucleus
6-hydroxydopamine on hypothalamic noradrenergic,
accumbens and striatum. Neuropsychopharmacology 26,
dopaminergic and 5-hydroxytryptaminergic neurons in
the male rat. Brain Research 365, 228–234.
Prisco S, Esposito E (1995). Differential effects of acute and
Martin-Ruiz R, Ugedo L, Honrubia MA, Mengod G,
chronic fluoxetine administration on the spontaneous
Artigas F (2001). Control of serotonergic neurons in rat
activity of dopaminergic neurones in the ventral tegmental
brain by dopaminergic receptors outside the dorsal raphe
area. British Journal of Pharmacology 116, 1923–1931.
nucleus. Journal of Neurochemistry 77, 762–775.
Prisco S, Pagannone S, Esposito E (1994). Serotonin-
Maskos U, Molles BE, Pons S, Besson M, Guiard BP,
dopamine interaction in the rat ventral tegmental area : an
Guilloux JP, Evrard A, Cazala P, Cormier A,
electrophysiological study in vivo. Journal of Pharmacology
Mameli-Engvall M, et al. (2005). Nicotine reinforcement
and Experimental Therapeutics 271, 83–90.
and cognition restored by targeted expression of nicotinic
Rapaport MH, Gharabawi GM, Canuso CM, Mahmoud RA,
Keller MB, Bossie CA, Turkoz I, Lasser RA, Loescher A,
McDonald WM, Richard IH, DeLong MR (2003).
Bouhours P, Dunbar F, Nemeroff CB (2006).
Prevalence, etiology, and treatment of depression
Effects of risperidone augmentation in patients with
in Parkinson’s disease. Biological Psychiatry 54, 363–375.
treatment-resistant depression : Results of open-label
Millan MJ, Lejeune F, Gobert A, Brocco M, Auclair A,
treatment followed by double-blind continuation.
Bosc C, Rivet JM, Lacoste JM, Cordi A, Dekeyne A (2000).
Neuropsychopharmacology 31, 2505–2513.
S18616, a highly potent spiroimidazoline agonist at
Rasmusson AM, Goldstein LE, Deutch AY, Bunney BS,
alpha(2)-adrenoceptors : II. Influence on monoaminergic
Roth RH (1994). 5-HT1a agonist+/x8-OH-DPAT
transmission, motor function, and anxiety in comparison
modulates basal and stress-induced changes in medial
with dexmedetomidine and clonidine. Journal
prefrontal cortical dopamine. Synapse 18, 218–224.
of Pharmacology and Experimental Therapeutics 295,
Reader TA, Briere R, Grondin L, Ferron A (1986). Effects of
p-chlorophenylalanine on cortical monoamines and on the
Nilsson LK, Schwieler L, Engberg G, Linderholm KR,
activity of noradrenergic neurons. Neurochemical Research
Erhardt S (2005). Activation of noradrenergic locus
Interactions between monoaminergic neurons
Reader TA, Gauthier P (1984). Catecholamines and serotonin
by the alpha-adrenergic agonist clonidine. Brain Research
in the rat central nervous system after 6-OHDA, 5-7-DHT
and p-CPA. Journal of Neural Transmission 59, 207–227.
Szabo ST, Blier P (2001). Effect of the selective noradrenergic
Reader TA (1982). Catecholamines and serotonin in rat
reuptake inhibitor reboxetine on the firing activity of
frontal cortex after PCPA and 6-OHDA : absolute amounts
noradrenaline and serotonin neurons. European Journal
and ratios. Brain Research Bulletin 8, 527–534.
Seager MA, Barth VN, Phebus LA, Rasmussen K (2005).
Szabo ST, Blier P (2002). Effects of serotonin
Chronic coadministration of olanzapine and fluoxetine
(5-hydroxytryptamine, 5-HT) reuptake inhibition plus
activates locus coeruleus neurons in rats : implications
5-HT(2A) receptor antagonism on the firing activity of
for bipolar disorder. Psychopharmacology (Berlin) 181,
norepinephrine neurons. Journal of Pharmacology and
Experimental Therapeutics 302, 983–991.
Seager MA, Huff KD, Barth VN, Phebus LA, Rasmussen K
Szabo ST, de Montigny C, Blier P (2000). Progressive
(2004). Fluoxetine administration potentiates the effect of
attenuation of the firing activity of locus coeruleus
olanzapine on locus coeruleus neuronal activity. Biological
noradrenergic neurons by sustained administration of
selective serotonin reuptake inhibitors. International Journal
Shelton RC, Williamson DJ, Corya SA, Sanger TM,
of Neuropsychopharmacology 3, 1–11.
Van Campen LE, Case M, Briggs SD, Tollefson GD
Tanda G, Carboni E, Frau R, Di Chiara G (1994). Increase of
(2005). Olanzapine/fluoxetine combination for
extracellular dopamine in the prefrontal cortex : a trait of
treatment-resistant depression : a controlled study of
drugs with antidepressant potential? Psychopharmacology
SSRI and nortriptyline resistance. Journal of Clinical
Ugedo L, Grenhoff J, Svensson TH (1989). Ritanserin, a
Shi WX, Pun CL, Zhang XX, Jones MD, Bunney BS (2000).
5-HT2 receptor antagonist, activates midbrain dopamine
Dual effects of D-amphetamine on dopamine neurons
neurons by blocking serotonergic inhibition.
mediated by dopamine and nondopamine receptors.
Psychopharmacology (Berlin) 98, 45–50.
Journal of Neuroscience 20, 3504–3511.
Ungless MA, Magill PJ, Bolam JP (2004). Uniform inhibition
Steffensen SC, Svingos AL, Pickel VM, Henriksen SJ (1998).
of dopamine neurons in the ventral tegmental area by
Electrophysiological characterization of GABAergic
aversive stimuli. Science 303, 2040–2042.
neurons in the ventral tegmental area. Journal of
White FJ, Wang RY (1984). Pharmacological characterization
of dopamine autoreceptors in the rat ventral tegmental
Svensson TH, Bunney BS, Aghajanian GK (1975). Inhibition
area : microiontophoretic studies. Journal of Pharmacology
of both noradrenergic and serotonergic neurons in brain
and Experimental Therapeutics 231, 275–280.
Het plassen en sproeien door katten in huis is zeer ongewenst gedrag en een zeer vervelend probleem voor hun eigenaren. Maar ook voor de katten zelf is het geen goede situatie omdat dit gedrag vaak voortkomt uit stress bij de kat. Gelukkig kunnen we het ontstaan van sproeigedrag vaak vermijden door de katten op een leeftijd van 6 maanden te castreren en steriliseren. Soms blijkt dit echter nie
74HC4075-Q100; 74HCT4075-Q100 Triple 3-input OR gate Rev. 1 — 22 May 2013 Product data sheet 1. General description The 74HC4075-Q100; 74HCT4075-Q100 is a triple 3-input OR gate. Inputs include clamp diodes. This enables the use of current limiting resistors to interface inputs to voltages in excess of VCC. This product has been qualified to the Automotive Electronics Council (
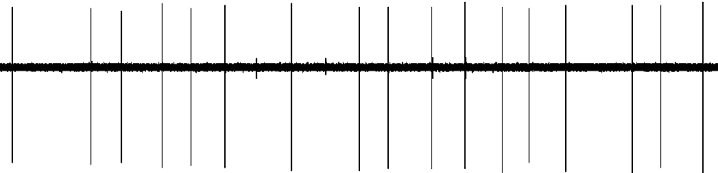
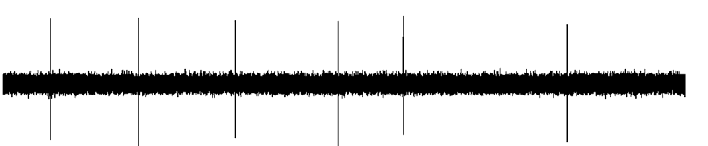
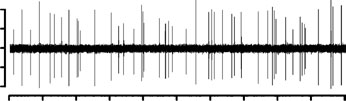
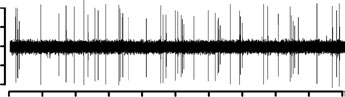 Figure 1. Effect of dopaminergic lesion with6-hydroxydopamine (6-OHDA) on the electrophysiologic
Figure 2. Effect of serotonergic lesion with 5,7-
activity of 5-HT neurons in the dorsal raphe (DR).
Figure 1. Effect of dopaminergic lesion with6-hydroxydopamine (6-OHDA) on the electrophysiologic
Figure 2. Effect of serotonergic lesion with 5,7-
activity of 5-HT neurons in the dorsal raphe (DR).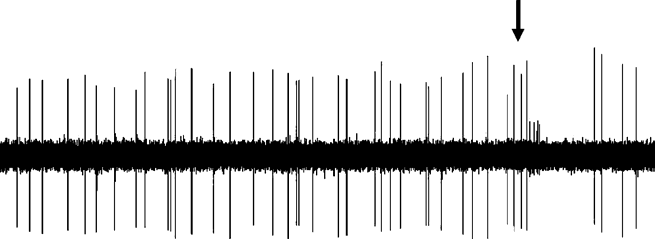
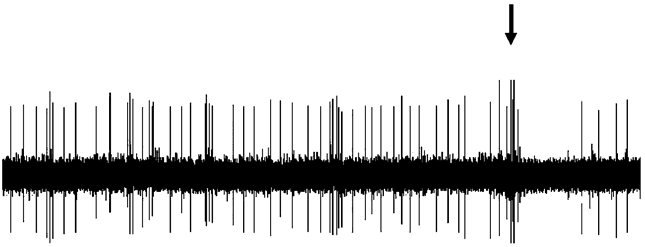
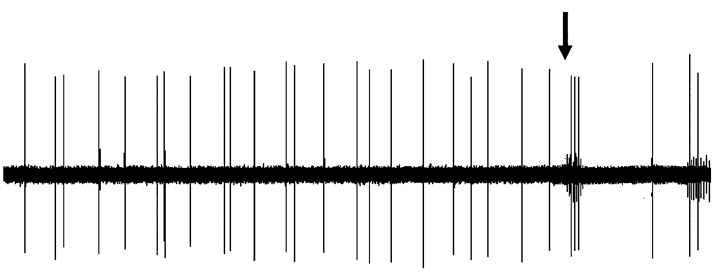
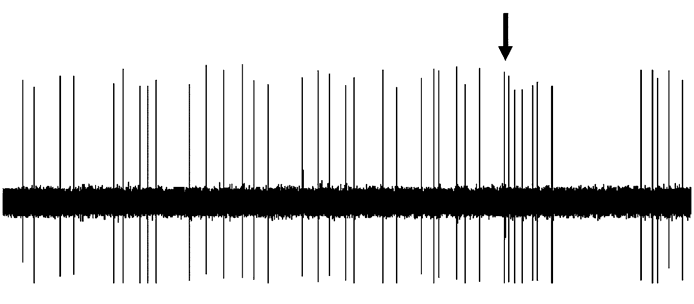 Figure 3. Effect of dopaminergic lesion with
6-hydroxydopamine (6-OHDA) on the electrophysiological
activity of locus coeruleus (LC) NE neurons discharging in asingle-spike mode. Examples of typical recordings of LC NE
Figure 4. Effect of dopaminergic lesion with
neurons obtained in (a) a sham-operated rat and (b) a ventral
6-hydroxydopamine (6-OHDA) on the electrophysiological
tegmental area (VTA) DA neuron-lesioned rat. The
activity of locus coeruleus (LC) NE neurons exhibiting a
characteristic increase in firing activity (without producing a
single-spike and burst activity. Examples of typical
burst) followed by a ‘ silence’ in response to pinch is
recordings of LC NE neurons obtained in (a) a sham-operated
indicated by an arrow. (c) Mean¡S.E.M. of frequency (Hz) of
rat and (b) a ventral tegmental area (VTA) DA neuron-
NE neurons recorded in the VTA of sham-operated rats
lesioned rat, each asterisk indicates a burst. The characteristic
(n=3) (grey histogram) and 6-OHDA-treated rats (n=7)
increase in firing activity followed by a ‘silence’ in response
(white histogram). * p<0.05 indicates significantly different
to pinch is indicated by an arrow. (c) Mean¡S.E.M. of
from sham-operated rats. The numbers within the histograms
frequency (Hz) of NE neurons. Number of single spikes/
indicate the number of neurons recorded.
Figure 3. Effect of dopaminergic lesion with
6-hydroxydopamine (6-OHDA) on the electrophysiological
activity of locus coeruleus (LC) NE neurons discharging in asingle-spike mode. Examples of typical recordings of LC NE
Figure 4. Effect of dopaminergic lesion with
neurons obtained in (a) a sham-operated rat and (b) a ventral
6-hydroxydopamine (6-OHDA) on the electrophysiological
tegmental area (VTA) DA neuron-lesioned rat. The
activity of locus coeruleus (LC) NE neurons exhibiting a
characteristic increase in firing activity (without producing a
single-spike and burst activity. Examples of typical
burst) followed by a ‘ silence’ in response to pinch is
recordings of LC NE neurons obtained in (a) a sham-operated
indicated by an arrow. (c) Mean¡S.E.M. of frequency (Hz) of
rat and (b) a ventral tegmental area (VTA) DA neuron-
NE neurons recorded in the VTA of sham-operated rats
lesioned rat, each asterisk indicates a burst. The characteristic
(n=3) (grey histogram) and 6-OHDA-treated rats (n=7)
increase in firing activity followed by a ‘silence’ in response
(white histogram). * p<0.05 indicates significantly different
to pinch is indicated by an arrow. (c) Mean¡S.E.M. of
from sham-operated rats. The numbers within the histograms
frequency (Hz) of NE neurons. Number of single spikes/
indicate the number of neurons recorded.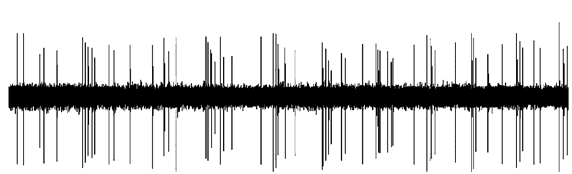
 Interactions between monoaminergic neurons
results indicate that the DA input exerts a direct excit-
atory effect, probably via D2 receptors on DR 5-HT
neurons while the influence of other non-dopaminergic
The i.c.v. injection of 5,7-DHT produced a robust
and selective decrease in brain 5-HT levels (Table 1)
leading to a 36 % increase in the discharge rate of VTA
DA neurons. This enhancement of DA neuronal ac-
tivity resulted from a higher number of bursts and
spikes per burst. The putative inhibitory effect of the
5-HT input on DA neurons suggested by the present
study is consistent with the finding that SSRIs, which
probably raise extracellular 5-HT levels in the VTA,
induce a slight decrease in the firing rate of VTA DAneurons (Di Mascio et al., 1998 ; Prisco and Esposito,
1995). The inhibitory influence of the 5-HT input on
DA neurons was further supported by the observation
that low doses of the 5-HT1A receptor agonist
8-OHDPAT, known to attenuate the electrical activity
of DR 5-HT neurons, increases the firing rate and the
number of burst discharge of DA neurons in the VTA
(Arborelius et al., 1993 ; Lejeune and Millan, 1998,
2000 ; Lejeune et al., 1997), and consequently DA
release at the somatodendritic (Chen and Reith, 1995)
and terminal levels (Ago et al., 2002 ; Arborelius et al.,
1993 ; Rasmusson et al., 1994 ; Tanda et al., 1994). How-
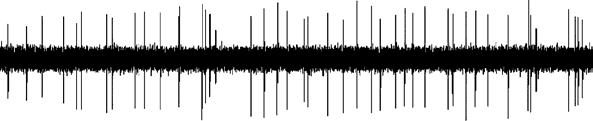
Figure 5. Effect of noradrenergic lesion with 6-
ever, high doses of 8-OHDPAT which preferentially
hydroxydopamine (6-OHDA) on the electrophysiological
activate post-synpatic receptors, also produce in-
activity of DA neurons in the ventral tegmental area (VTA).
Interactions between monoaminergic neurons
results indicate that the DA input exerts a direct excit-
atory effect, probably via D2 receptors on DR 5-HT
neurons while the influence of other non-dopaminergic
The i.c.v. injection of 5,7-DHT produced a robust
and selective decrease in brain 5-HT levels (Table 1)
leading to a 36 % increase in the discharge rate of VTA
DA neurons. This enhancement of DA neuronal ac-
tivity resulted from a higher number of bursts and
spikes per burst. The putative inhibitory effect of the
5-HT input on DA neurons suggested by the present
study is consistent with the finding that SSRIs, which
probably raise extracellular 5-HT levels in the VTA,
induce a slight decrease in the firing rate of VTA DAneurons (Di Mascio et al., 1998 ; Prisco and Esposito,
1995). The inhibitory influence of the 5-HT input on
DA neurons was further supported by the observation
that low doses of the 5-HT1A receptor agonist
8-OHDPAT, known to attenuate the electrical activity
of DR 5-HT neurons, increases the firing rate and the
number of burst discharge of DA neurons in the VTA
(Arborelius et al., 1993 ; Lejeune and Millan, 1998,
2000 ; Lejeune et al., 1997), and consequently DA
release at the somatodendritic (Chen and Reith, 1995)
and terminal levels (Ago et al., 2002 ; Arborelius et al.,
1993 ; Rasmusson et al., 1994 ; Tanda et al., 1994). How-
Figure 5. Effect of noradrenergic lesion with 6-
ever, high doses of 8-OHDPAT which preferentially
hydroxydopamine (6-OHDA) on the electrophysiological
activate post-synpatic receptors, also produce in-
activity of DA neurons in the ventral tegmental area (VTA).