Do you want to buy antibiotics online without prescription? https://buyantibiotics24h.net/ - This is pharmacy online for you!
Changes in stimulus salience as a result of stimulus preexposure: evidence from aversive and appetitive testing procedures
Learning & Behavior2003, 31 (2), 185-191
Changes in stimulus salience as a result of
stimulus preexposure: Evidence from aversive
In two experiments, rats received preexposure to three compound flavor stimuli, AX, BX, and CX,
where X represents a saline solution. AX and BX were presented in alternation; CX, on a separate block
of trials. The value of X was then modified, being devalued by aversive conditioning in Experiment 1,
and rendered valuable by the induction of a state of salt need in Experiment 2. When given a choice be-
tween BX and CX, the rats consumed more of BX than of CX in Experiment 1, and more of CX than of
BX in Experiment 2, suggesting that B and C differed in their ability to modulate the response governed
by the X element. It was suggested that blocked preexposure to CX reduces the salience of the C stim-
ulus but that the salience of B is maintained by preexposure in which BX is alternated with AX. The im-
plications of this result for the phenomenon of perceptual learning are discussed.
Our standard theories of classical conditioning (see,
salience can be explained in terms of known associative
e.g., Wagner, 1981) assume the existence of a represen-
tational node for each of the events that can be shown to
The experimental evidence on which Hall’s (2003) ac-
be effective as a conditioned or unconditioned stimulus
count is based comes, in part, from a recent series of
(CS or US). Learning is held to occur when two nodes
studies reported by Blair and Hall (2003) examining gen-
are activated concurrently and to consist of changes in
eralization in flavor aversion learning after preexposure
the strength of associative links between them. Since the
to the stimuli. Our basic procedure made use of three fla-
degree of change will depend on the extent to which each
vors, A, B, and C, that were rendered similar by the ad-
of the nodes is activated, it is important to be able to
dition of an explicit element (X) common to each. (Here
specify the relationship between the external stimulus
A represents a lemon solution; B and C represent sucrose
and the level of activity that its presentation induces in its
and saline solutions, counterbalanced; and X, a solution
node. The usual assumption has been that the level of ac-
of quinine.) One of these compounds (AX) was trained
tivity in a node will be determined directly by the inten-
as a CS in the flavor-aversion paradigm; the aim of the
sity (or salience) of the stimulus that is applied. But Hall
experiment was to see how different schedules of pre-
(2003) has challenged this assumption, arguing, on the
exposure to the stimuli would influence generalization
basis of evidence from experiments on perceptual learn-
from AX to the test compounds BX and CX. In the pre-
ing, that the ability of a given stimulus to activate its node
exposure phase, there were four presentations of each of
can change with experience—that mere exposure to a
the three stimuli. Presentations of CX occurred in one
stimulus can engage a learning process that produces block of trials; presentations of AX and BX, however,
a change in the effective salience of the stimulus. Hall
were intermixed, occurring on alternate trials. It was
suggests that repeated exposure to a stimulus will nor-
found (Blair & Hall, 2003, Experiment 1) that the aver-
mally result in a loss of salience but that exposure to in-
sion established to AX generalized less readily to BX
termixed (alternating) presentations of two similar stim-
than to CX. In explaining these results, we argued that
uli will maintain, or even enhance, the effective salience
the aversion displayed on the test trials in this procedure
of their distinctive features. The experiments to be de-
would be largely determined by the ability of the condi-
scribed in this article were designed to explore the latter
tioned element, X, to evoke its conditioned response
suggestion, and, in particular, to assess whether the evi-
(CR) and that the differential responding shown to BX
dence that has been interpreted as indicating changes in
and CX indicated that the B and C differed in their ability
to interfere with the expression of this CR. We suggested
that the effective salience of B and C had been modified
This work was supported by a studentship from the Biotechnology and
during the preexposure phase—that the salience of C had
Biological Science Research Council to Chris Blair. We thank C. Bonardi,
declined during the block of exposure trials with CX but
E. Mondragón, and M. Symonds for helpful discussion. Correspondence
that alternating trials with AX and BX had maintained or
concerning this article should be addressed to either C. A. J. Blair or
G. Hall, who are both at the Department of Psychology, University of
enhanced the salience of their unique features (and in
York, York YO10 5DD, England (e-mail: GH1@york.ac.uk).
particular of B). The less salient C stimulus would be less
Copyright 2003 Psychonomic Society, Inc.
likely to interfere with the ability of the conditioned el-
compound as in the previous experiment) was used as
ement X to evoke its CR than would the more salient B
the CS. When tested with BX and CX, the animals again
stimulus, producing the result obtained.
consumed less of CX than of BX. This is the result to be
The interpretation offered by Hall (2003) implies the
expected if the test performance depends on a difference
existence of a novel (or, at least, nonassociative) learn-
between B and C in their ability to interfere with the ex-
ing process that modulates the perceptual effectiveness
pression of the CR governed by X; the more salient B
of stimuli. But before we commit ourselves to this con-
will be more effective in this regard than the less salient
clusion, it is important to be sure that the effects on
C, both when X has been conditioned in compound with
which it is based cannot be explained in terms of stan-
A, as in the previous experiment, and when X has been
dard associative mechanisms. And, as Hall (2003) has
conditioned alone. McLaren and Mackintosh (2000),
acknowledged, the associative account of perceptual
however, predict that there should be no difference on
learning proposed by McLaren and Mackintosh (2000;
test when X has been conditioned alone. According to
see also McLaren, Kaye, & Mackintosh, 1989) can sup-
their account, BX differs from CX in that B is able to in-
ply an explanation for the results reported by Blair and
hibit the A representation whereas C is not; but this dif-
ference will be relevant only when the conditioning pro-
McLaren and Mackintosh (2000) point out that expo-
cedure is one that has endowed A with some associative
sure to a compound stimulus will result in the formation
strength (as when conditioning is given with AX); it
of excitatory associations among the elements of the
should not affect performance when conditioning has
compound (Rescorla & Cunningham, 1978). The pre-
exposure procedure used by Blair and Hall (2003) can
Given that the results of Blair and Hall’s (2003) Ex-
thus be expected to establish associations between A and
periment 5 appear to have theoretically important impli-
X, B and X, and C and X. That between A and X could
cations, we attempted, in the experiments to be reported
contribute to the magnitude of the CR observed on the
here, to confirm and extend them. In the present Experi-
test trial with CX. Conditioning with AX as the CS will
ment 1, we adopted essentially the same design and pro-
endow both A and X with associative strength, and the
cedure, but we changed the nature of the flavors used; in
aversion shown to CX on the test will no doubt be deter-
particular, we used saline rather than quinine as the X el-
mined largely by the strength of the conditioned X ele-
ement. To anticipate, we succeeded in replicating the re-
ment. But although A is not presented on the test, its
sult obtained by Blair and Hall in their Experiment 5, thus
strength could still play a role, because the existence of
extending the generality of the effect. More important,
the A–X association would allow the X element of the
the outcome of Experiment 1 formed the basis for a fur-
test stimulus to contact the US representation by way of
ther investigation of the phenomenon. According to the
the chain X–A–US. Such a process can be expected to
interpretation offered by Hall (2003), the difference be-
operate when CX is the test stimulus, but when BX is the
tween BX and CX on test derives from a difference in the
test stimulus another factor must be taken into account.
ability of B and C to interfere with the responding gov-
According to standard associative theory, preexposure
erned by the conditioned X element. In the experiments
consisting of alternating trials with AX and BX will not
described so far, X has been subjected to aversive condi-
only establish excitatory associations between the ele-
tioning so that the effect of presenting the salient B ele-
ments of each compound, but also result in the formation
ment along with X has been to attenuate the suppression
of inhibitory associations between their unique features.
of consumption generated by X. But if Hall’s (2003) in-
In particular, once the excitatory, within-compound,
terpretation is correct, B should be more effective than C
A–X association has been established, presentation of
in attenuating any response controlled by the X ele-
BX will activate the representation of the absent A ele-
ment—if, for instance, X is particularly valued by the rat,
ment (by way of the X–A association), a circumstance
then the presence of B in the BX compound can be ex-
that, according to Wagner (1981), will result in the for-
pected to lead to a reduction in the amount consumed
mation of inhibitory links between B and A and between
(compared with the amount of CX consumed).
X and A. The excitatory link between X and A will be
Using saline as the X element allows a test of this pre-
reestablished on the next AX trial, but B, being a unique
diction. Injecting rats with a mixture of furosemide and
predictor of the absence of A, will acquire net inhibitory
deoxycorticosterone acetate (Furo-Doca) induces a state
strength. As a consequence, on test trials with BX, acti-
of salt need and renders them eager to consume a saline
vation of A by way of the X–A link will be inhibited by
solution (Cruz, Perelle, & Wolf, 1977). Animals in this
the presence of B, and the associative strength acquired
state should readily consume compounds such as BX
by A will not be able to contribute to the CR.
and CX, which contain saline. But if alternating pre-
In an attempt to test the validity of this account, we
exposure renders B relatively salient (with respect to the
conducted a further study (Blair & Hall, 2003, Experi-
C element preexposed in a block of CX trials), then they
ment 5) in which, as before, rats received exposure con-
will be less ready to consume the BX compound (in
sisting of alternating presentations of AX and BX and a
which the presence of B will interfere with the percep-
separate block of CX trials. They then received aversion
tion of X) than the CX compound. This prediction was
conditioning in which the X element (rather than the AX
EXPERIMENT 1
were given unrestricted access to water on both drinking sessions. The second conditioning trial, given in the morning session of the
In this experiment, all subjects (rats) received an ini-
next day, was identical to the first and was followed by a further re-
tial preexposure phase consisting of trials with three
covery day. The third conditioning trial was identical to the second.
compound flavor stimuli, AX, BX, and CX. A, B, and C
Water was again available in the afternoon session following this
conditioning trial, and 1 further recovery day preceded the test
were commercially produced food flavorings; X was a
saline solution. This was followed by aversion condi-
On the following morning session, subjects were given a 30-min
tioning with X as the CS. All then received a two-bottle
choice test, receiving access to 30 ml of BX and to 30 ml of CX in
test in which the compound BX and CX were presented.
two separate drinking tubes presented simultaneously. The two
(In the experiment by Blair & Hall, 2003, on which this
tubes were inserted into the cage on either side of the aperture used
study was based, the test procedure consisted of a series
for presentations of the single tube given during earlier stages of
training. The two spouts were separated by a distance of 5 cm. The
of single-bottle tests. The use of a two-bottle test in the
position of the tubes was counterbalanced in such a way that half
present experiment was dictated by the desire to match
the rats were presented with BX on the right, and half with CX on
the procedure with that to be used in Experiment 2, in
the right. Water was made available for 30 min in the afternoon ses-
which the use of a two-bottle test was dictated by the par-
sion. Over the next 4 days, four further test sessions were given, the
ticular procedures employed.) If this procedure should
procedure being identical to that just described.
generate an effect equivalent to that demonstrated by
Blair and Hall (2003, Experiment 5), one would expect
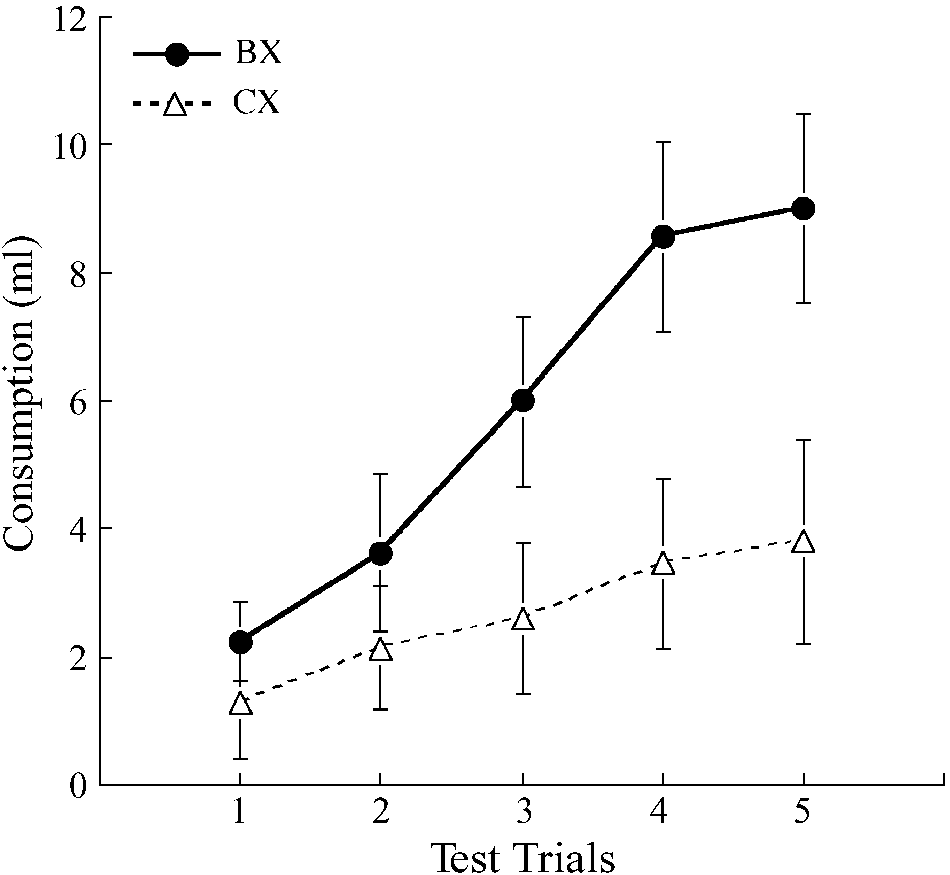
Results and Discussion
to find that the subjects would consume more of BX than
The rats consumed all of the fluid that was made avail-
able on the preexposure sessions. The conditioning pro-
cedure successfully established an aversion to X. Group
mean consumption was 9.31 ml on Trial 1, 9.32 ml on
Subjects and Apparatus. The subjects were 8 experimentally
Trial 2, and 4.08 ml on Trial 3. All subjects showed a re-
naive male hooded Lister rats with a mean ad-lib weight of 367 g at
duction in consumption from Trial 1 to Trial 3.
the start of the experiment. The rats were housed singly, with con-
Group mean consumption scores for the test sessions
tinuous access to food, in a colony room that was artificially lit
are presented in Figure 1. The rats consumed rather lit-
from 8:00 a.m. to 8:00 p.m. each day. Access to water was restricted
tle of either solution on the first test session, but con-
as detailed below. Presentations of the various solutions used as thestimuli in these experiments were given in the home cages.
sumption increased over sessions, presumably as a result
The solutions used as experimental stimuli were administered at
of extinction of the aversion conditioned to X. The sub-
room temperature in 50-ml plastic centrifuge tubes equipped with
jects consumed more of BX than of CX on each of the
a rubber stopper to which was fitted a stainless steel, ball-bearing-
five tests, with the difference between the means for con-
tipped spout. The following solutions were used: A compound con-sisting of 0.08 M saline (NaCl) and almond (2% v/v almond flavor-ing supplied by Supercook, Leeds, U.K.); a compound of 0.08 M
saline and vanilla (1% v/v Supercook vanilla flavoring) sulfate; anda compound of 0.08 M saline and peppermint (0.5% v/v Supercook
peppermint flavoring). Consumption was measured by weighingthe tubes before and after trials, to the nearest 0.1 g. The US for the
conditioning trials was an intraperitoneal injection of 0.3 M lithiumchloride (LiCl) at 10 ml/kg of body weight. Procedure. A schedule of water deprivation was initiated by re-
moving the standard water bottles overnight. On each of the fol-lowing 3 days, access to water was restricted to two daily sessions
of 30 min, at 11:00 a.m. and 5:00 p.m. Presentation of fluids con-tinued to be given at these times throughout the experiment.
Over the next 6 days (the preexposure phase), all subjects received
four presentations of each of the three compound flavors AX, BX,and CX. Half the animals were first given 4 days of intermixed ac-
cess to flavors AX and BX, with 10 ml of AX being presented dur-ing the first daily drinking session and 10 ml of BX during the sec-
ond. This was followed by 2 days of blocked presentations of CX,in which 10 ml of this flavor was made available in both morningand afternoon drinking sessions. The remainder of the subjects re-
ceived the blocked presentations of CX on the first 2 days of thephase followed by 4 days of AX and BX. For all animals, flavor A
was peppermint and flavor X was saline. The critical test flavors, Band C, were counterbalanced with half the animals receiving vanilla
Figure 1. Experiment 1: Mean consumption of flavor com-
as B and almond as C, and half receiving the reverse arrangement. pounds BX and CX over five two-bottle test trials. The rats had
Three conditioning trials followed. The first was given in the
all received preexposure consisting of a block of trials with CX
morning session the day after preexposure ended. It consisted of a
and alternating trials with AX and BX, prior to aversion condi-
30-min presentation of 9 ml of X, followed immediately by an in-
tioning with X. Error bars represent within-subjects standard
jection of LiCl. The rats were given free access to water in the af-
error, computed on scores adjusted for variation between sub-
ternoon session. The next day was a recovery day on which animals
jects (Bakeman & McArthur, 1996).
sumption of the two flavors increasing throughout. This
saline. We gave rats preexposure consisting of alternating
description of the results was confirmed by an analysis
trials with AX and BX and a separate block of CX trials.
of variance (ANOVA) conducted on the data summa-
We then gave them an injection of Furo-Doca followed
rized in the figure, with test stimulus (BX or CX) and
by a test session in which BX and CX were presented. We
trial as the factors. There was no significant main effect
expected that the rats in a state of salt need would tend to
of test stimulus [F(1,7) = 2.18; here and elsewhere a sig-
consume both of these compounds readily, to the extent
nificance level of p < .05 was adopted], but there was a
that they were able to perceive the saline component. The
significant effect of trial [F(4,28) = 16.32], and, criti-
question of interest was whether the supposedly more
cally, a significant interaction between trial and stimulus
salient B element would interfere with this tendency more
[F(4,28) = 4.54]. Pairwise comparisons using Tukey’s
than the less salient C element would, resulting in a
test revealed that the scores for the two flavors differed
greater consumption of CX than of BX.
It will be noted that, in this experiment, the looked-for
The data presented in the figure pool the results for
pattern on the test session (more consumption of CX
two subgroups, one presented with CX on the first block
than of BX) was the opposite of that predicted (and
of preexposure trials, the other presented with CX on the
found) in Experiment 1. To find this pattern would allow
second block of preexposure trials. With only 4 animals
us to rule out some possible, but theoretically uninter-
in each of these subgroups, our ability to detect any dif-
esting, explanations for the result of that experiment.
ference between them will be limited. There was some
One of these was the possibility that the preexposure
indication, however, that the effect of interest (the low
schedules used in these experiments were differentially
level of consumption of CX on test) was more substan-
effective in their ability to produce habituation. If habit-
tial in the subgroup given CX in the second block. Pool-
uation occurs more readily with the alternating schedule
ing over all test trials showed that this group drank a
than with the blocked arrangement, then the BX compound
mean of 5.7 ml of BX and 1.6 ml of CX; the equivalent
would be less likely to evoke neophobia than would the
scores for the group given CX first were 6.0 ml and
CX compound, resulting in greater consumption of BX
3.8 ml. This difference between the subgroups was not,
than CX—the result obtained in Experiment 1. But this
however, statistically reliable. An ANOVA paralleling
account cannot predict greater consumption of CX than
that just described, but including subgroup as a variable,
of BX, the result anticipated in this experiment.
yielded neither a significant main effect of this variable
An alternative interpretation of the results of Experi-
[F(1,6) = 1.49] nor any significant interaction—for the
ment 1 can be derived from the fact that in that experiment,
subgroup 3 stimulus interaction, F < 1; for the triple
presentations of BX were always given during afternoon
sessions in preexposure, whereas CX also occurred dur-
Although this experiment made use of different fla-
ing morning sessions (but see Blair & Hall, 2003, Ex-
vors as the stimuli and a different test procedure, its re-
periment 1B). Aversion conditioning to X occurred dur-
sults were entirely consistent with those reported by
ing a morning session. Accordingly, if time of day can
Blair and Hall (2003, Experiment 5). Rats that had re-
serve as a cue that mediates generalization, it might be
ceived aversion conditioning with flavor X drank more
argued that generalization would be greater to CX than
of it on test when it was presented in compound with B
to BX, producing the effect seen in Experiment 1. But
than when it was presented in compound with C. This
this argument does not apply to the present experiment—
outcome is thus in accord with the suggestion that the
given that the procedure does not involve a conditioning
schedule of preexposure used in the first phase of the ex-
phase, there is no basis on which the time-of-day factor
periment rendered stimulus B effectively more salient
can play a role and therefore no reason to predict any dif-
than stimulus C and thus better able to disrupt the ex-
ference in consumption between CX and BX on the test.
pression of the aversion governed by X.
Finally, it could be argued that the results of Experi-
ment 1, in which the difference between BX and CX de-
EXPERIMENT 2
veloped only over the course of several nonreinforced
test sessions, might be a consequence of a difference be-
Our proposal that the results of Experiment 1 reflect
tween the two test stimuli in the rate at which extinction
differences in salience between B and C depends on the
occurs. Given that the aversion shown on test will be
(surely plausible) assumption that a more salient stimulus
largely a consequence of the associative strength gov-
will be more effective than a less salient stimulus in in-
erned by a stimulus element, X, that is common to both
terfering with the response elicited by some other stimu-
test compounds, it is not clear how such a difference
lus with which it is compounded. In Experiment 1, this
might arise. Nonetheless, it is worth noting that differ-
response was the aversion to X established by an aversive
ential extinction could not easily explain the anticipated
conditioning procedure in which X served as the CS. But
results of the present experiment, in which the test con-
if our interpretation is correct, B and C should differ in
sisted of a single presentation of BX and CX.
their ability to modulate any response controlled by X. In
the present experiment, we made use of the fact that X
was a solution of saline and that an injection of Furo-
The subjects were 16 experimentally naive male hooded Lister
Doca would increase the readiness of rats to consume
rats with a mean ad-lib weight of 352 g at the start of the experi-
ment. The experiment was run in two replications, each using 8 an-
ference between the groups was quite marked. The ef-
imals. In the first replication, 1 animal became ill prior to the test
fect shown in the figure was almost entirely a conse-
phase so that data were available for 15 subjects. The flavors used
quence of the behavior of the subgroup (n = 8) given CX
as stimuli were those described for Experiment 1. The treatment
first. This group consumed 22.6 ml of CX and 7.1 ml of
used to induce a sodium appetite was a subcutaneous injection of
0.5 ml of a mixture of 10 mg furosemide (Furo) and 5 mg of de-
BX. The subgroup given CX second (n = 7) consumed
oxycorticosterone acetate (Doca) dispersed in 20 ml of distilled
rather less overall, and the difference between the scores
for BX (11.2 ml) and CX (11.9 ml) was negligible. An
The subjects initially received exposure to AX, BX, and CX, the
ANOVA conducted on these data revealed a significant
procedure being identical to that described for the preexposure
main effect of test stimulus [BX or CX; F(1,13) = 6.25]
phase of Experiment 1. One hour after the end of the final pre-
and no significant main effect of subgroup [F(1,13) =
exposure session (the afternoon session on Day 6 of preexposure),
all animals received an injection of Furo-Doca. The food was then
3.76], but there was a significant interaction between
removed from the home cages in the colony room, and the subjects
these variables [F(1,13) = 5.24]. Analysis of simple ef-
were given free access to distilled water overnight. On the follow-
fects showed that the scores for BX and CX differed sig-
ing day, the distilled water was removed from the cages 3 h prior to
nificantly in the subgroup given CX first [F(1,13) =
the test session (the morning drinking session). On test, the subjects
12.29], but not in the subgroup given CX second (F < 1).
were given a free-access choice test, receiving 30 ml of flavor BX
The theoretical implications of this difference be-
and an identical volume of flavor CX, in the same manner as in Ex-
tween the subgroups will be taken up in the General Dis-
periment 1. Only one such test was given (the effects of the Furo-
cussion. For the time being, we will simply note that the
overall pattern of the results (as shown in Figure 2) ac-
Results and Discussion
cords with the interpretation that we offered for the re-
The rats consumed all of the fluid made available on
sults of Experiment 1. In that experiment, the stimulus
each of preexposure sessions, with no evidence of neo-
element X was made aversive and the animals consumed
less of CX than of BX on test, a result we interpreted as
The results of the test session, group means for con-
showing that B was more effective than C in modulating
sumption of BX and CX, are presented in Figure 2. It
the response controlled by X. In this experiment, stimu-
shows that subjects consumed more of CX than of BX.
lus element X was made desirable by the administration
A within-subjects, one-way ANOVA confirmed that the
of Furo-Doca just after preexposure. The resulting salt
difference between the two test stimuli was statistically
need meant that when given a choice, subjects were mo-
tivated to consume the flavor compound that they per-
As in the previous experiment, we looked separately at
ceived as being richer in salt. Our prediction, therefore,
the performance of the two counterbalanced subgroups,
was that the perceptually salient B element would be
which were given CX presentations as the first or as the
more likely to interfere with perception of the desirable
second block of preexposure trials. In this case, the dif-
salt than would the less salient C element, leading to
greater consumption of CX than of BX—the result that
As we pointed out in the introduction to this experi-
ment, the finding of greater consumption of CX than of
BX in the present experiment rules out some, relatively
trivial, explanations that might be offered for the results
of Experiment 1. It also allows us to address a possible
explanation that can be derived from an elaboration of
the theory proposed by McLaren et al. (1989). Accord-
ing to that theory, it will be recalled, the preexposure
schedule employed in these experiments will result in the
formation of excitatory links between X and each of A,
B, and C; it will also allow the formation of inhibitory
links between A and B. When animals are conditioned
with X (as in Experiment 1), the excitatory X– C link
should generate activity in the representation of stimulus
C; but the existence of inhibitory A– B links might limit
the extent to which the representations of these stimuli
can be activated. Finally, if it is accepted that the asso-
Figure 2. Experiment 2: Mean consumption of flavor com-
ciatively activated representation of a stimulus can un-
pounds BX and CX on the choice test. The rats had all received
dergo conditioning (see Hall, 1996), it follows that C
preexposure consisting of a block of trials with CX and alternat-
will be more likely than A or B to acquire associative
ing trials with AX and BX, followed by the induction of a salt need
strength during reinforced X trials. Differences in the ac-
immediately prior to the test. Error bars represent within-subjects standard error, computed on scores adjusted for variation be-
quisition of strength by C could then explain why the an-
tween subjects (Bakeman & McArthur, 1996).
imals showed a greater aversion to CX than to BX in Ex-
periment 1. Resting, as it does, on a number of debatable
exposure phase of these experiments will also undergo
assumptions, this explanation may seem implausible, but
habituation and thus become less effective at eliciting
there is nothing in the results of Experiment 1 to rule it
whatever unobserved covert response they would nor-
out entirely. It cannot apply to the results of the present
mally evoke in the animal; it is this loss of effectiveness
experiment, however, since the mechanism proposed de-
that we have equated with a change in stimulus salience.
pends critically on events occurring during the condi-
The real problem is to explain why this loss of effective-
tioning trials with X. The central feature of this experi-
ness should be attenuated or reversed for A and B when
ment was that the value of X was modified by means of
preexposure consists of alternating trials with AX and
a procedure that did not involve presentations of X.
According to Gibson’s (1969) account of perceptual
GENERAL DISCUSSION
learning, exposure to similar stimuli will engage a dif-ferentiation process that will enhance the perceptual ef-
Taken together with the results reported by Blair and
fectiveness of unique stimulus features. Differentiation
Hall (2003), the experiments described here allow the
is held to occur more readily when the subject has a
following conclusions. Preexposure to the compound
chance to compare the relevant stimuli. It might be sug-
stimuli AX, BX, and CX will, among other things, mod-
gested, then, that alternating preexposure is particularly
ify the properties of their unique features, A, B, and C.
effective in maintaining or strengthening the effective-
When the preexposure consists of alternating trials with
ness of A and B because these stimuli are presented ac-
AX and BX, and a separate block of CX trials, the con-
cording to a schedule that promotes stimulus compari-
sequence appears to be that the effective salience of the
son. The obvious problem with this suggestion is that
B element (and presumably also of the A element, al-
although AX and BX were indeed presented in alterna-
though this was not tested directly) is greater than that of
tion in these experiments, the interval between succes-
the C element. This difference is evidenced by the effects
sive trials was at least 5 h, making it unlikely that any ac-
of compounding B and C with another element (X). When
tivation produced by presentation of the first stimulus
X is aversive, compounding it with B results in less of an
would still be ongoing when the second stimulus oc-
aversion than does compounding it with C. When X has
curred. What does seem possible, however, is that the
a positive value, compounding it with B results in less
central representation of each of the critical stimulus fea-
being consumed than when it is compounded with C.
tures will be activated associatively on each trial (after
This pattern of results is parsimoniously explained in
the first) in the alternating schedule. Alternating trials
terms of the proposal that the more salient B is more
with AX and BX can be expected to establish and main-
likely to interfere with the ability of X to evoke its re-
tain excitatory connections between X and A and be-
tween X and B, allowing A to be activated associatively
The proposal that alternating exposure to two similar
on BX trials and B to be activated associatively on AX
stimuli will enhance the salience of their unique ele-
trials. Hall (2003) has advanced the hypothesis that the
ments (or at least results in a lesser loss than is produced
associative activation of a stimulus representation in the
by repeated presentation of just one stimulus) has the ca-
absence of direct presentation of the stimulus itself will
pability to explain a wide range of perceptual learning
act to reverse the loss of salience that occurs when the
effects. The central feature of these effects (for a recent
review, see Hall, 2001) is that appropriately scheduled
Comparison of the counterbalanced subgroups of the
exposure to similar stimuli can increase the ease with
present experiments (those given CX preexposure in the
which they can subsequently be discriminated. The
first block vs. those given CX preexposure in the second
essence of discrimination learning is that behavior must
block) allows a test of this hypothesis. For both sub-
come to be controlled by the unique, distinguishing fea-
groups, the alternating schedule used for presentations
tures of the stimuli rather than by features they hold in
of AX and BX ensures that associative activation of the
common. A learning process that enhances the (relative)
representation of B will occur regularly on AX trials.
salience of the unique features of similar stimuli would
According to the hypothesis, the salience of B should
thus be capable of generating the observed effects.
therefore be maintained or enhanced in both. The fate of
It remains to explain the nature of the learning process
the C element will, however, differ in the two subgroups.
responsible for changes in stimulus salience, and here we
When CX is presented second during preexposure, there
have little more than speculation to offer. That repeated
will be no opportunity for the associative activation of C
exposure to a given stimulus (such as CX in the present
in the absence of the stimulus, and its decline in salience
experiments) might cause a loss of salience (in both C
should proceed unhindered. But when CX is presented
and X) is not problematic. The phenomenon of habitua-
first, the formation of the within-compound C–X asso-
tion demonstrates that repeated presentation of a stimu-
ciation during the first block of trials will mean that C
lus that initially evokes an overt response will reduce the
can be activated associatively on the subsequent AX and
effectiveness of the stimulus in this respect. Although
BX trials (at least until the point at which these trials re-
they do not evoke any obvious overt response, it is rea-
sult in extinction of the X– C association). C should
sonable to assume that the stimuli presented in the pre-
therefore lose salience less readily in the latter condition.
It follows that the difference in effectiveness between B
sults are challenging for the hypothesis offered by Hall
and C on the test should be greater in the subgroup that
(2003), but, importantly for our present concern, the
received CX during the second block of preexposure tri-
basic pattern obtained is not to be easily explained by es-
als than in the subgroup that received CX in the first
tablished associative theories of perceptual learning as
Unfortunately, the results of the present experiments
are equivocal on this issue. In Experiment 1, the means
REFERENCES
for the two subgroups were entirely in accord with our
prediction; the subgroup given CX first in preexposure
Bakeman, R., & McArthur, D. (1996). Picturing repeated measures:
Comments on Loftus, Morrison, and others. Behavior Research
drank more of CX on the test than did the subgroup given
Methods, Instruments, & Computers, 28, 584-589.
CX second, implying that the salience of C was greater
Blair, C. A. J., & Hall, G. (2003). Perceptual learning in flavor aver-
in the former group than in the latter. That the difference
sion: Evidence for learned changes in stimulus effectivenes
between the subgroups was small (and not statistically
Cruz, C. E., Perelle, I. B., & Wolf, G. (1977). Methodological as-
reliable) would also be expected, given that the CX-first
arrangement supplies conditions that are less than opti-
mal for the associative activation of C (and thus for pro-
Gibson, E. J. (1969). Principles of perceptual learning and develop-
ducing the increase in effective salience envisaged by
ment. New York: Appleton-Century-Crofts.
our interpretation). In this subgroup, C may be activated
Hall, G. (1996). Learning about associatively activated stimulus rep-
resentations: Implications for acquired equivalence and perceptual
associatively during the initial presentations of AX and
BX that follow exposure to CX, but, in the absence of
Hall, G. (2001). Perceptual learning: Association and differentiation.
further CX trials, it is to be expected that extinction of
In R. R. Mowrer & S. B. Klein (Eds.), Handbook of contemporary
the within-compound association between X and C will
learning theories (pp. 367-407). Mahwah, NJ: Erlbaum.
Hall, G. (2003). Learned changes in the sensitivity of stimulus repre-
limit this effect just to these initial trials. This would ex-
sentations: Associative and nonassociative mechanisms.
plain why consumption of CX was only slightly higher in
the subgroup for which CX was presented first, and why,
McLaren, I. P. L., Kaye, H., & Mackintosh, N. J. (1989). An asso-
overall, the means indicate a clear perceptual learning
ciative theory of the representation of stimuli: Applications to per-
effect (significantly higher consumption of BX than CX)
ceptual learning and latent inhibition. In R. G. M. Morris (Ed.), Par-allel distributed processing: Implications for psychology andneurobiology (pp. 102-130). Oxford: Oxford University Press,
The results for the equivalent analysis of Experiment 2
are more problematic. Here the subgroup given CX first
McLaren, I. P. L., & Mackintosh, N. J. (2000). An elemental model
(but not that given CX second) showed a profound dif-
of associative learning: I. Latent inhibition and perceptual learning. Animal Learning & Behavior, 28, 211-246.
ference in consumption of the two test solutions—the re-
Rescorla, R. A., & Cunningham, C. L. (1978). Within-compound fla-
verse of the effect predicted (and obtained in Experi-
ment 1). We can see no obvious explanation for this
outcome, and we will need to carry out further work to
Wagner, A. R. (1981). SOP: A model of automatic memory processing
confirm its reliability. However this may turn out, it is
in animal behavior. In N. E. Spear & R. R. Miller (Eds.), Informationprocessing in animals: Memory mechanisms (pp. 5-47). Hillsdale,
encouraging that the overall findings of Experiment 2
are largely in accord with conclusions drawn from a
body of work that has, until now, relied on the condi-
tioned flavor aversion procedure. Some aspects of the re-
revision accepted for publication January 22, 2003.)
SAFETY DATA SHEET Emergency Phone: 1800-033-882 (24 hrs) +61 3 9663 2130 (24 hrs) Dow AgroSciences Australia Ltd. Frenchs Forest NSW 2086 PROFUMETM GAS FUMIGANT 1. PRODUCT AND COMPANY IDENTIFICATION: PRODUCT: ProFumeTM Gas Fumigant In case of accident or if you feel unwell, seek medical advice immediately (show the label PURPOSE: Fumigant for insect control.
Centre for Health Informatics, University of New South Wales, and Department of Health Science and Technology, Aalborg Universityy Telemedicine - Aid to decision-making in healthcarey The amount of information available to answer questions Æ The use of keyword-based search methods to locate the answersy These methods tend to overload with a lot of irrelevant Æ The precision in the search r
 EXPERIMENT 1
EXPERIMENT 1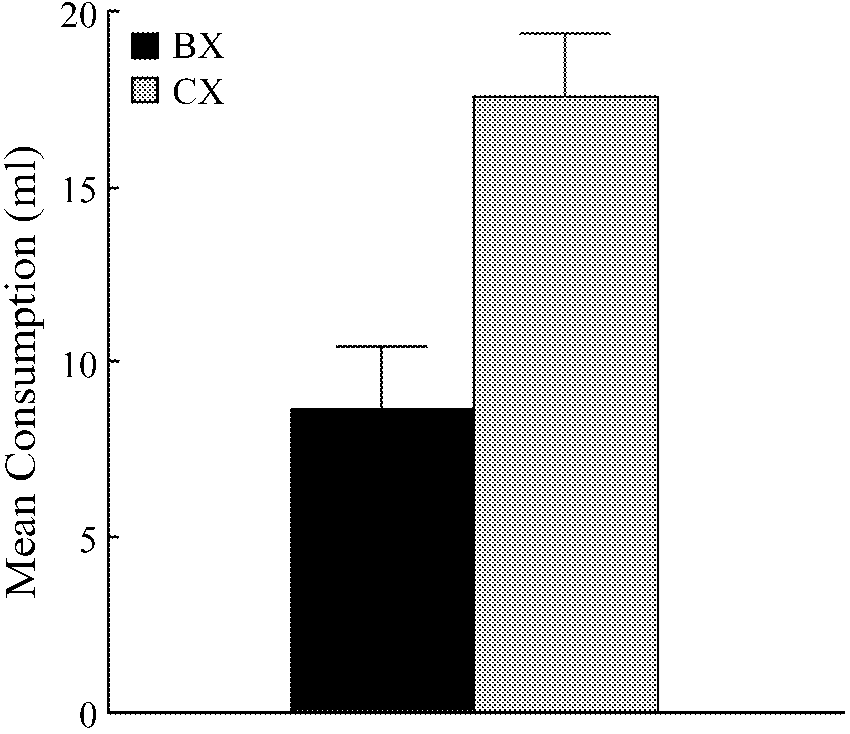 ment. The experiment was run in two replications, each using 8 an-
ference between the groups was quite marked. The ef-
imals. In the first replication, 1 animal became ill prior to the test
fect shown in the figure was almost entirely a conse-
phase so that data were available for 15 subjects. The flavors used
quence of the behavior of the subgroup (n = 8) given CX
as stimuli were those described for Experiment 1. The treatment
first. This group consumed 22.6 ml of CX and 7.1 ml of
used to induce a sodium appetite was a subcutaneous injection of
0.5 ml of a mixture of 10 mg furosemide (Furo) and 5 mg of de-
BX. The subgroup given CX second (n = 7) consumed
oxycorticosterone acetate (Doca) dispersed in 20 ml of distilled
rather less overall, and the difference between the scores
for BX (11.2 ml) and CX (11.9 ml) was negligible. An
The subjects initially received exposure to AX, BX, and CX, the
ANOVA conducted on these data revealed a significant
procedure being identical to that described for the preexposure
main effect of test stimulus [BX or CX; F(1,13) = 6.25]
phase of Experiment 1. One hour after the end of the final pre-
and no significant main effect of subgroup [F(1,13) =
exposure session (the afternoon session on Day 6 of preexposure),
all animals received an injection of Furo-Doca. The food was then
3.76], but there was a significant interaction between
removed from the home cages in the colony room, and the subjects
these variables [F(1,13) = 5.24]. Analysis of simple ef-
were given free access to distilled water overnight. On the follow-
fects showed that the scores for BX and CX differed sig-
ing day, the distilled water was removed from the cages 3 h prior to
nificantly in the subgroup given CX first [F(1,13) =
the test session (the morning drinking session). On test, the subjects
12.29], but not in the subgroup given CX second (F < 1).
ment. The experiment was run in two replications, each using 8 an-
ference between the groups was quite marked. The ef-
imals. In the first replication, 1 animal became ill prior to the test
fect shown in the figure was almost entirely a conse-
phase so that data were available for 15 subjects. The flavors used
quence of the behavior of the subgroup (n = 8) given CX
as stimuli were those described for Experiment 1. The treatment
first. This group consumed 22.6 ml of CX and 7.1 ml of
used to induce a sodium appetite was a subcutaneous injection of
0.5 ml of a mixture of 10 mg furosemide (Furo) and 5 mg of de-
BX. The subgroup given CX second (n = 7) consumed
oxycorticosterone acetate (Doca) dispersed in 20 ml of distilled
rather less overall, and the difference between the scores
for BX (11.2 ml) and CX (11.9 ml) was negligible. An
The subjects initially received exposure to AX, BX, and CX, the
ANOVA conducted on these data revealed a significant
procedure being identical to that described for the preexposure
main effect of test stimulus [BX or CX; F(1,13) = 6.25]
phase of Experiment 1. One hour after the end of the final pre-
and no significant main effect of subgroup [F(1,13) =
exposure session (the afternoon session on Day 6 of preexposure),
all animals received an injection of Furo-Doca. The food was then
3.76], but there was a significant interaction between
removed from the home cages in the colony room, and the subjects
these variables [F(1,13) = 5.24]. Analysis of simple ef-
were given free access to distilled water overnight. On the follow-
fects showed that the scores for BX and CX differed sig-
ing day, the distilled water was removed from the cages 3 h prior to
nificantly in the subgroup given CX first [F(1,13) =
the test session (the morning drinking session). On test, the subjects
12.29], but not in the subgroup given CX second (F < 1).