Do you want to buy antibiotics online without prescription? https://buyantibiotics24h.net/ - This is pharmacy online for you!
Doi:10.1016/s1095-6433(03)00108-9
Comparative Biochemistry and Physiology Part A 135 (2003) 425–433
Effects of inhibition gastric acid secretion on arterial acid–base
status during digestion in the toad Bufo marinus
Johnnie B. Andersena,*, Denis V. Andradeb, Tobias Wanga
of Zoophysiology, Institute of Biological Sciences, University of Aarhus, Universitetsparken Building 131,
bDepartment de Zoologia, Universidade Estadual Paulista, Rio Claro SP 13506-900, Brazil
Received 18 December 2002; received in revised form 8 April 2003; accepted 9 April 2003
Abstract
Digestion affects acid–base status, because the net transfer of HCl from the blood to the stomach lumen leads to an
HCO3 levels in both extra- and intracellular compartments. The increase in plasma w
tide, is particularly pronounced in amphibians and reptiles, but is not associated with an increased arterial pH, becauseof a concomitant rise in arterial PCO2 caused by a relative hypoventilation. In this study, we investigate whether thepostprandial increase in PaCO2 of the toad Bufo marinus represents a compensatory response to the increased plasmaw
or a state-dependent change in the control of pulmonary ventilation. To this end, we successfully prevented the
alkaline tide, by inhibiting gastric acid secretion with omeprazole, and compared the response to that of untreated toadsdetermined in our laboratory during the same period. In addition, we used vascular infusions of bicarbonate to mimicthe alkaline tide in fasting animals. Omeprazole did not affect blood gases, acid–base and haematological parameters infasting toads, but abolished the postprandial increase in plasma w
and the rise in arterial PCO2 that normally
peaks 48 h into the digestive period. Vascular infusion of
HCO3 , that mimicked the postprandial rise in plasma
HCO3 , led to a progressive respiratory compensation of arterial pH through increased arterial PCO2. Thus, irrespective
of whether the metabolic alkalosis is caused by gastric acid secretion in response to a meal or experimental infusion ofbicarbonate, arterial pH is being maintained by an increased arterial PCO2. It seems, therefore, that the elevated PCO2,occuring during the postprandial period, constitutes of a regulated response to maintain pH rather than a state-dependentchange in ventilatory control.
ᮊ 2003 Elsevier Science Inc. All rights reserved. Keywords: Toad; B. marinus; Digestion; Acid–base status; Alkaline tide; Gastric acid secretion; Omeprazole; Ventilatory control
1. Introduction
. Digestion causes a rise in metabolicrate, the ‘specific dynamic action of food’ (SDA),
Many ectothermic vertebrates eat large meals at
infrequent intervals and the ensuing digestion is
presence of food in the stomach stimulate a net
HCl secretion from the blood to the stomach lumenthat leads to an increase in
HCO3 , the so-called ‘alkaline tide’, is
1095-6433/03/$ - see front matter ᮊ 2003 Elsevier Science Inc. All rights reserved. doi:10.1016/S1095-6433(03)00108-9
J.B. Andersen et al. / Comparative Biochemistry and Physiology Part A 135 (2003) 425–433
pronounced in amphibians and reptiles, but the
immersion into a 1.0 g ly1 benzocaine solution
(ethyl p-amino benzoate Sigma᭨ E 1501), and
surgery started when the corneal reflex disap-
peared. The right femoral artery was occlusively
cannulated through an incision in the leg, and the
acterised, therefore, by a metabolic alkalosis that
catheter was secured to the back of the animal by
is almost fully compensated by a respiratory aci-
three or four sutures. The surgery normally lasted
dosis apparently caused by a relative hypoventila-
less than 30 min and all toads regained normal
righting reflexes within 30 min after being placed
under running tap water. All toads were treated
qualitatively similar respiratory compensations of
with enrofloxacin (Baytril; 2 mg kgy1, i.p.) to
the alkaline tide have been observed in mammals
prevent infections. When the toads had regained
normal reflexes, each individual animal was trans-
ferred to an experimental chamber (40=30=20
the regulation of ventilation during digestion is
cm) containing wet paper towels and a dry area.
geared at maintaining pH rather than PCO2. How-
These containers were maintained within a climatic
ever, because digestion is associated with large
chamber at a constant temperature of 25 8C, the
metabolic increments, it is possible that the rise in
arterial PCO2 (PaCO2) simply reflects an ineffective
ventilatory compensation to the increased meta-
bolic rate, leading to an un-regulated maintenanceof pH. Alternatively, it is possible that the
increased PaCO2 during digestion is caused byinduction of a more relaxed state with low respon-
2.2.1. Effects of omeprazole on blood gas compo-
siveness to ventilatory stimuli during the postpran-
To inhibit gastric acid secretion, omeprazole was
To study whether toads regulate pHa or PaCO2
given orally to six fasting toads prior to the
during digestion, we measured acid–base parame-
experiments. Omeprazole was dissolved in meth-
ters of animals, where gastric acid secretion was
ylcellulose (1.5%) and administrated through a
inhibited by the specific proton-pump inhibitor
soft rubber tubing inserted into the stomach
omeprazole. Omeprazole has been previously
through the mouth. A dose of 0.06 mg kgy1 (2
shown to uncouple Hq and Cly secretion in the
ml of 28 mg kgy1 pr kg toad) omeprazole was
gastric mucosa in the frog Rana catesbeiana
applied daily over 4 days before cannulation, and
a final dose was administered a few hours before
formed on the marine toad (Bufo marinus), which
has been extensively studied with regards to its
A blood sample from fasting animals was with-
acid–base regulation and from which we have data
drawn 24 h after surgery, as we have previously
shown that arterial blood gases and acid–base
parameters of B. marinus have stabilised at thistime , and analysed
2. Materials and methods
immediately (see below). Then, the animals wereforce-fed rat pups amounting to 7.0"0.3% of body
mass. Subsequent blood samples were taken 24and 48 h after feeding.
Toads, Bufo marinus (Linnaeus, 1758) of unde-
A group of un-treated toads, where blood sam-
termined sex and body masses between 230–522
ples were taken at the same time into the digestive
g (355"24 g, mean"1 S.E.M.) were obtained
period, were included for comparison. These data
from Lemberger (Oshkosh, WI, USA) and kept at
the University of Aarhus for several months. The
toads were kept at 23–28 8C in large containers
during the same period as those described in the
with access to running water and dry areas and
present study and using the same batch of toads
fed mealworms daily. Toads were anaesthetised by
J.B. Andersen et al. / Comparative Biochemistry and Physiology Part A 135 (2003) 425–4332.2.2. Effects of bicarbonate injection on arterial
response compared to untreated animals. The SDA
response of untreated B. marinus was used from a
A control blood sample was withdrawn 24 h
previous study using the same batch of animals
after surgery. Then, bicarbonate was injected as a
1.5 to 3.5 ml bolus, depending on the mass of the
animal, of 1 mol ly1 NaHCO3 giving a final
bonate injection, a one-way ANOVA for repeated
concentration of 6.9"0.04 mmol kgy1 toad. Blood
measures was employed. All differences among
samples were withdrawn 1, 2, 6, 12 and 24 h after
means were assessed by a SNK post-hoc test. The
level of significance was chosen at the P-0.05level. All data in text and figures are presented as
3. Blood gas analysis
Arterial blood was analysed for oxygen tension
4. Results
(PaO2), pH, haematocrit, blood haemoglobin con-centration (wHb x
4.1. Arterial acid–base status during digestion
carbon dioxide content of plasma (wCO x). PaO
and pHa were measured with Radiometer (Copen-hagen, Denmark) electrodes maintained in a BMS
In the untreated control toads, digestion was
3 electrode set-up at 25 8C while displaying the
associated with a 12 mmol ly1 increase in plasma
output on a Radiometer PHM 73. Haematocrit was
determined in duplicate as the fractional red cell
volume after centrifugation (12 000 rpm for 3
so that arterial pH did not change during
conversion to cyanmethaemoglobin, applying a
millimolar extinction coefficient of 10.99 at 540
Arterial acid–base status of fasting animals was
not significantly affected by omeprazole treatment
affected. Thus, in the omeprazole-treated toads,
change significantly during digestion when com-
bers were maintained at 40 8C. Haemoglobin
bound oxygen (HbO2) was calculated as wO x
remained relatively constant and significantly low-
er than in the untreated toads throughout the
digestive period . The difference between
omeprazole-treated and untreated animals becomes
(HbO2sat) was calculated as: HbO2ywHbx, under
even more apparent when depicting arterial acid–
the assumption that all Hb was functional. Arterial
base parameters in a Davenport diagram .
carbon dioxide tension (PaCO2) was calculated
The omeprazole-treated animals show a minor
respiratory disturbance, whereas the untreated ani-
mals show a metabolic alkalosis, which is com-
pensated by a respiratory acidosis, thereby keeping
Assuming that the carbonate concentration
Digestion was not associated with changes in
blood oxygen levels and haematological parame-
4.2. Acid–base status after vascular injections ofbicarbonate
Significant effects of digestion were found by
the use of a one-way ANOVA for repeated meas-
Vascular injection of bicarbonate caused a sig-
ures. A two-way ANOVA was employed to iden-
tify significant effects of omeprazole on the SDA
26.5"1.37 to 36.0"1.77 mmol ly1 1 h after
J.B. Andersen et al. / Comparative Biochemistry and Physiology Part A 135 (2003) 425–433
was associated with a substantial increase in pHafrom 7.74"0.03 to 7.91"0.02, at unchangedPaCO .
2 However, as seen in the Davenport diagram
, the metabolic alkalosis was progressivelycompensated by an increased PaCO2, (i.e. a respi-ratory acidosis). Plasma w
icantly elevated 24 h after the infusion, while pHawas fully compensated. The bicarbonate injectionhad no effect on blood oxygen levels and haema-tological parameters . 5. Discussion
Our aim of this study was to investigate whether
the increased PaCO2 during digestion in Buforepresents a compensatory response to increasedplasma w
HCO3 , (i.e. the alkaline tide) or whether
the increased PaCO2 represents a state-dependentchange in the control of pulmonary ventilation. Tothis end, we successfully prevented the alkalinetide by inhibiting gastric acid secretion with ome-prazole and, in addition, vascular infusions ofbicarbonate mimicked the alkaline tide in fastinganimals. 5.1. Effects of digestion on arterial blood gases inB. marinus
In untreated toads, digestion was associated with
of the simultaneous increase in PaCO2, pHa didnot change during digestion. A similar respiratorycompensation of the postprandial metabolic alka-losis, (i.e. the alkaline tide) has been observed inall amphibians and reptiles where blood sampleshave been obtained from undisturbed animalsusing indwelling catheters
Fig. 1. Arterial acid–base parameters in the toad Bufo marinusbefore and during digestion. The circles show the response of
untreated control animals, while the response of omeprazole-
treated toads is shown by the squares. Fasting values are pre-
marinus and other amphibians and reptiles is
sented as open symbols. (a) arterial pH; (b) plasma
numerically larger than in mammals. This is par-
tially due to the smaller meal size ingested by
tension, PaCO2. Means that are different from the fasting valueare marked with an asterisk, while significant effects of ome-
mammals and a consequence of a more regular
prazole treatment are marked with a dagger. Data are presented
feeding pattern where gastric acid secretion is
as mean "1 S.E.M. (Ns6 in each group).
continuously countered by pancreatic base secre-tion to the small intestine. Furthermore, the mam-
infusion . There was no significant reduc-
malian kidney responds effectively to metabolic
tion during the next 24 h. This increase in plasma
acid–base disturbances and the alkaline tide is
rapidly reduced by increased base output in the
to changes elicited by digestion in untreated toads
case in amphibians where transport of acid–base
J.B. Andersen et al. / Comparative Biochemistry and Physiology Part A 135 (2003) 425–433
Fig. 2. Davenport diagram showing plasma w
and arterial pH during fasting and digestion in the toad Bufo marinus. Animals
treated with omeprazole are shown with the squares and untreated control animals are shown by circles. The Davenport diagram includestwo in vitro non-bicarbonate buffer lines (dotted lines, bNB) determined by and isobars for the partial pressureof CO2 in arterial blood (PaCO2, curved lines). Data are presented as mean "1 S.E.M. (Ns6 in each group).
relevant ions over the bladder and kidney is less
5.2. Acid–base regulation after inhibition of gas-
effective than the mammalian kidney (see
tric acid secretion with omeprazole
. Inthe present experiments this is revealed by the
mechanism, which is the final step in the secretory
infusion: less than half of the extra bicarbonate,
process of the ATP-driven proton pump, and inhib-
present in the plasma 1 h after infusion, had been
its both basal and meal-stimulated secretion of
removed 24 h into the experiment . In R.catesbeiana, transepithelial acid–base exchange is
. Arterial blood gases and haematological
parameters of fasting omeprazole-treated toads
were not significantly different from untreated
Table 1Effects of digestion on arterial blood gases and haematological parameters in omeprazole-treated and untreated toads (Bufo marinus)
2 ), oxygen tension (PaO2), haemoglobin oxygen saturation (HbO2sat), haematocrit (Hct), haemoglobin
4 ), mean cellular haemoglobin concentration, plasma pH, plasma carbon dioxide (PaCO2 ) and bicarbonate concen-
Values are mean "1 S.E.M (Ns6 in each group)
J.B. Andersen et al. / Comparative Biochemistry and Physiology Part A 135 (2003) 425–433
remains uncertain whether acid–base status offasting animals is affected by omeprazole. Inmammals, omeprazole is considered to be veryspecific and without side effects (e.g. and arterial acid–base parameters of fastingrats are not affected by omeprazole (Wang, Norlenand Haakanson, unpublished). Almost half of theomeprazole-treated toads vomited within 48 h afterforce feeding, and while their blood gas composi-tion did not differ from those completing digestion(data not shown), we excluded these animals fromthe study. It is likely that inhibition of gastric acidsecretion impaired the digestive ability and stimu-lated the emetic reflex, and secondary adverseeffects of omeprazole cannot be ruled out.
in omeprazole-treated toads, which indicates aneffective inhibition of the proton pump of theparietal cells in the gastric mucosa. The inhibitionof the alkaline tide by omeprazole is consistentwith the postprandial increase in plasma w
being caused by a rise in plasma strong iondifference, as protons and chloride are secretedinto the stomach lumen. Omeprazole also inhibitedthe postprandial rise in plasma w
The inhibition of the postprandial rise in plasma
respiratory acidosis reflects a ventilatory compen-sation to maintain pHa. 48 h into the postprandialperiod, omeprazole had completely abolished theincreased PaCO2. This indicates that the relativehypoventilation during the postprandial period is aregulated response that act to maintain pHa bymodulating PaCO2. A similar conclusion wasreached in experiments on B. constrictor, whereomeprazole fully abolished the increase in PaCO2
Fig. 3. Effects of a bicarbonate injection at 0h (6.9"0.04
mmol kgy1) on arterial acid–base parameters in the toad Bufo
tilatory regulation of pHa, rather than PaCO2, is
marinus. Open symbol denotes pre-injected, whereas closed
further supported by the observation that vascular
symbols denotes post-injected animals. (a) arterial pH; (b)
bicarbonate infusion led to an increased PaCO
that re-established pHa at the control level 24 h
bon dioxide tension, PaCO2. Significant differences from thepre-injected values are marked with an asterisk. Data are pre-
after infusion, however, the response at 24 h after
sented as mean "1 S.E.M. (Ns6).
feeding was less clear, because there was a tenden-cy, albeit not statistically significant, for an
toads and were similar to previous studies on B.
Our study cannot reveal, which chemoreceptors
are involved in mediating ventilatory regulation of
ever, appeared slightly higher in omeprazole-treat-
pHa during the postprandial period, but it indicates
ed animals, which was also observed in the snake
that the overall modality of the chemoreceptors
controlling ventilation, at fast and during digestion,
given the lack of statistically significant effects, it
is pHa and not PaCO2. The ventilatory response to
J.B. Andersen et al. / Comparative Biochemistry and Physiology Part A 135 (2003) 425–433
Fig. 4. Davenport diagram showing plasma w
and arterial pH of fasting Bufo marinus before (open symbols) and after (closed
symbols) a vascular injection of bicarbonate (6.9"0.04 mmol kgy1). The Davenport diagram includes two in vitro non-bicarbonatebuffer lines (dotted lines, bNB) determined by and isobars for the partial pressure of CO2 in arterial blood(PaCO2, curved lines). Data are presented as mean "1 S.E.M. (Ns6).
hypercapnia (the combination of increased PCO2
and reduced pH) of B. marinus is primarily driven
permeability of the blood brain barrier to ions and
by central chemoreceptors in the medulla
CO2 has not been characterised in ectothermic
vertebrates, but it is likely that the slow rate for
central receptors are responsible for the postpran-
the development of the alkaline tide allows for the
dial response, it is required that metabolic acid–
metabolic alkalosis to be transmitted from the
base disturbances are transmitted from plasma to
blood to the CSF. This may even be the case in
the cerebrospinal fluid (CSF). In mammals, the
mammals, since the small alkaline tide is associ-
blood brain barrier separating blood from CSF, is
ated with small respiratory compensations
rather impermeable to ions while changes in
PCO2 are readily transmitted between the two
Table 2Effects of a bicarbonate injection (6.9"0.04 mmol kgy1) on arterial blood gases and haematological parameters in toads (Bufo marinus)
2 ), oxygen tension (PaO2), haemoglobin oxygen saturation (HbO2sat), haematocrit (Hct), haemoglobin
4 ), mean cellular haemoglobin concentration, plasma pH, plasma carbon dioxide (PaCO2 ) and bicarbonate concen-
Values are mean "1 S.E.M (Ns6)
J.B. Andersen et al. / Comparative Biochemistry and Physiology Part A 135 (2003) 425–433
relevant ions between plasma and CSF could also
American alligator, Alligator mississippiensis. J. Exp. Biol.
explain the rather slow and progressive ventilatory
compensation to the alkalosis following infusion
Cameron, J.N., 1971. Rapid method for determination of total
carbon dioxide in small blood samples. J. Appl. Physiol.
of bicarbonate in the present study. It may also be
important to consider pHyPCO2 sensitivity of the
Christoforides, C., Hedley-Whyte, J., 1969. Effect of temper-
ature and hemoglobin concentration on solubility of O2 in
Thus, while the mechanisms underlying the
blood. J. Appl. Physiol. 27 (5), 592–596.
postprandial regulation of pHa remains to be
Erdt, H., 1915. Die Tagesschwankungen der Kohlensaur
understood in more detail, the present study adds
nung der Alveorluft und ihre Ursachen. Deut. Arch. Klin. Med. 117, 497–516.
strong support for the view that the relative hypo-
Fencl, V., 1986. Acid–base balance in cerebrospinal fluid. In:
ventilation during the elevated metabolic rate asso-
Fishman, A.P., Cherniak, N.S., Widdicombe, J.G., Geiger,
ciated with digestion, reflects a regulation of arte-
S.R. (Eds.), Handbook of Physiology, section 3. The respi-
rial pH rather than a state dependent change in
ratory system, volume II. Control of breathing, part 1. APS,
Glass, M.L., Wood, S.C., Hoyt, R.W., Johansen, K., 1979.
Chemical control of breathing in the lizard, Varanus exan-Acknowledgments thematicus. Comp. Biochem. Physiol. A 62, 999–1003.
Hicks, J.W., Wang, T., Bennett, A.F., 2000. Patterns of cardi-
ovascular and ventilatory response to elevated metabolicstates in the lizard, Varanus exanthematicus. J. Exp. Biol.
Research Council and FAPESP. We are grateful to
Johannes Overgaard and Steve Warburton for help
Higgins, H.L., 1914. The influence of food, posture, and other
factors on the alveolar carbon dioxide tension in man. Am. J. Physiol. 34, 14–126. References
Niv, Y., Fraser, G.M., 2002. The alkaline tide phenomenon. J.
Ou, L.C., Tenney, S.M., 1974. Post-prandial rise in alveolar
Andersen, J.B., Jensen, F.B., Wang, T., 2001. Effects of
CO and ventilatory response in cats. Respir. Physiol. 22,
temperature and oxygen availability on circulating catecho-
lamines in the toad Bufo marinus. Comp. Biochem. Physiol.
Overgaard, J., Busk, M., Hicks, J.W., Jensen, F.B., Wang, T.,
1999. Acid–base status and arterial oxygen transport follow-
Andersen, J.B., Wang, T., 2002. Effects of anaesthesia on
ing feeding in the snake Python molurus. Comp. Biochem.
blood gases, acid–base status and ions in the toad Bufomarinus. Comp. Biochem. Physiol. 131A, 639–646.
Sachs, G., Shin, J.M., Briving, C., Wallmark, B., Hersey, S.,
Andersen, J.B., Wang, T., 2003. Cardio-respiratory conse-
1995. The pharmacology of the gastric acid pump: the H ,
quences of forced activity and digestion in toads. Physiol.
Kq ATPase. Annu. Rev. Pharmacol. Toxicol. 35, 277–305.
Andrade, D.V., De Toledo, L.F, Abe, A.S., Wang, T. Ventilatory
Secor, S.M., Hicks, J.W., Bennett, A.F., 2000. Ventilatory and
compensation of the alkaline tide during digestion in the
cardiovascular responses of a python (Python molurus) to
snake Boa constrictor. (in review).
exercise and digestion. J. Exp. Biol. 203, 2447–2454.
Boutilier, R.G., Randall, D.J., Shelton, G., Toews, D.P., 1979.
Smatresk, N.J., Smits, A.W., 1991. Effects of central and
Acid–base relationship in the blood of the toad, Bufo
peripheral chemoreceptor stimulation on ventilation in the
marinus. I: The effect of environmental CO
marine toad, Bufo marinus. Resp. Physiol. 83, 223–238.
Starlinger, M.J., Hollands, M.J., Rowe, P-H., Matthews, J.B.,
Branco, L.G., Glass, M.L., Hoffmann, A., 1992. Central
Silen, W., 1986. Chloride transport of frog gastric fundus:
chemoreceptor drive to breathing in unanesthetized toads.
effects of omeprazole. Am. J. Physiol. 250, G118–G126. Bufo paracnemis. Resp. Physiol. 87 (2), 195–204.
Toews, D.P., Stiffler, D.F., 1989. The role of the amphibian
Branco, L.G., Glass, M.L., Wang, T., Hoffmann, A., 1993.
kidney and bladder in the regulation of acid–base relevant
Temperature and central chemoreceptor drive to ventilation
ions. Can. J. Physiol. 67, 3064–3069.
in toad (Bufo paracnemis). Resp. Physiol. 93 (3), 337–346.
Tucker, V.A., 1967. Method for oxygen content and dissocia-
Bridges, C.R., Bicudo, J.E.P.W., Lykkeboe, G., 1979. Oxygen
tion curves on microliter blood samples. J. Appl. Physiol.
content measurement on blood containing haemocyanin.
Comp. Biochem. Physiol. A 62, 457–462.
Tufts, B.L., Toews, D.P., 1986. Renal function and acid–base
Busk, M., Jensen, F.B., Wang, T., 2000a. The effects of feeding
balance in the toad Bufo marinus during short-term dehy-
on blood gases, acid–base parameters and selected metab-
dration. Can. J. Zool. 64, 1054–1057.
olites in the bullfrog Rana catesbeiana. Am. J. Physiol.
Van Slyke, D.D., Stillman, E., Cullen, G.E., 1917. Studies of
acidosis. V. Alveolar carbon dioxide and plasma bicarbonate
Busk, M., Overgaard, J., Hicks, J.W., Bennett, A.F., Wang, T.,
in normal men during digestive rest and activity. J. Biol.
2000b. Effects of feeding on arterial blood gases in the
J.B. Andersen et al. / Comparative Biochemistry and Physiology Part A 135 (2003) 425–433
Wang, T., Burggren, W.W., Nobrega, E., 1995. Metabolic,
Wood, S.C., Malvin, G.M., 1991. Physiological significance
ventilatory, and acid–base responses associated with specific
of behavioral hypothermia in hypoxic toads (Bufo marinus).
dynamic action in the toad Bufo marinus. Physiol. Zool. 68,
Zijlstra, W.G., Buursma, A., Zwart, A., 1983. Molar absorptiv-
Wang, T., Busk, M., Overgaard, J., 2001. The respiratory
ities of human hemoglobin in the visible spectral range. J.
consequences of feeding in amphibians and reptiles. Comp.
Correspondence Infant mortality (per thousand livebirths) group on HIV in women and children. Mortality among infected and uninfectedAfrica: a pooled analysis. Lancet 2004; 364: Abrams EJ, Wiener J, Carter R, et al. Maternalhealth factors and early pediatric antiretroviraltherapy influence the rate of perinatal HIV-Idisease progression in children. AIDS , 2003; Table: Infant m
SECRETARIA DE SALUD ACUERDO que establece los Lineamientos para la adquisición de medicamentos asociados al Catálogo Universal de Servicios de Salud por las entidades federativas con recursos del Sistema de Protección Social en Salud. Al margen un sello con el Escudo Nacional, que dice: Estados Unidos Mexicanos.- Secretaría de Salud. JOSE ANGEL CORDOVA VILLALOBOS, Secretario de Salud
 Comparative Biochemistry and Physiology Part A 135 (2003) 425–433
Effects of inhibition gastric acid secretion on arterial acid–base
status during digestion in the toad Bufo marinus
Johnnie B. Andersena,*, Denis V. Andradeb, Tobias Wanga
of Zoophysiology, Institute of Biological Sciences, University of Aarhus, Universitetsparken Building 131,
bDepartment de Zoologia, Universidade Estadual Paulista, Rio Claro SP 13506-900, Brazil
Received 18 December 2002; received in revised form 8 April 2003; accepted 9 April 2003
Abstract
Comparative Biochemistry and Physiology Part A 135 (2003) 425–433
Effects of inhibition gastric acid secretion on arterial acid–base
status during digestion in the toad Bufo marinus
Johnnie B. Andersena,*, Denis V. Andradeb, Tobias Wanga
of Zoophysiology, Institute of Biological Sciences, University of Aarhus, Universitetsparken Building 131,
bDepartment de Zoologia, Universidade Estadual Paulista, Rio Claro SP 13506-900, Brazil
Received 18 December 2002; received in revised form 8 April 2003; accepted 9 April 2003
Abstract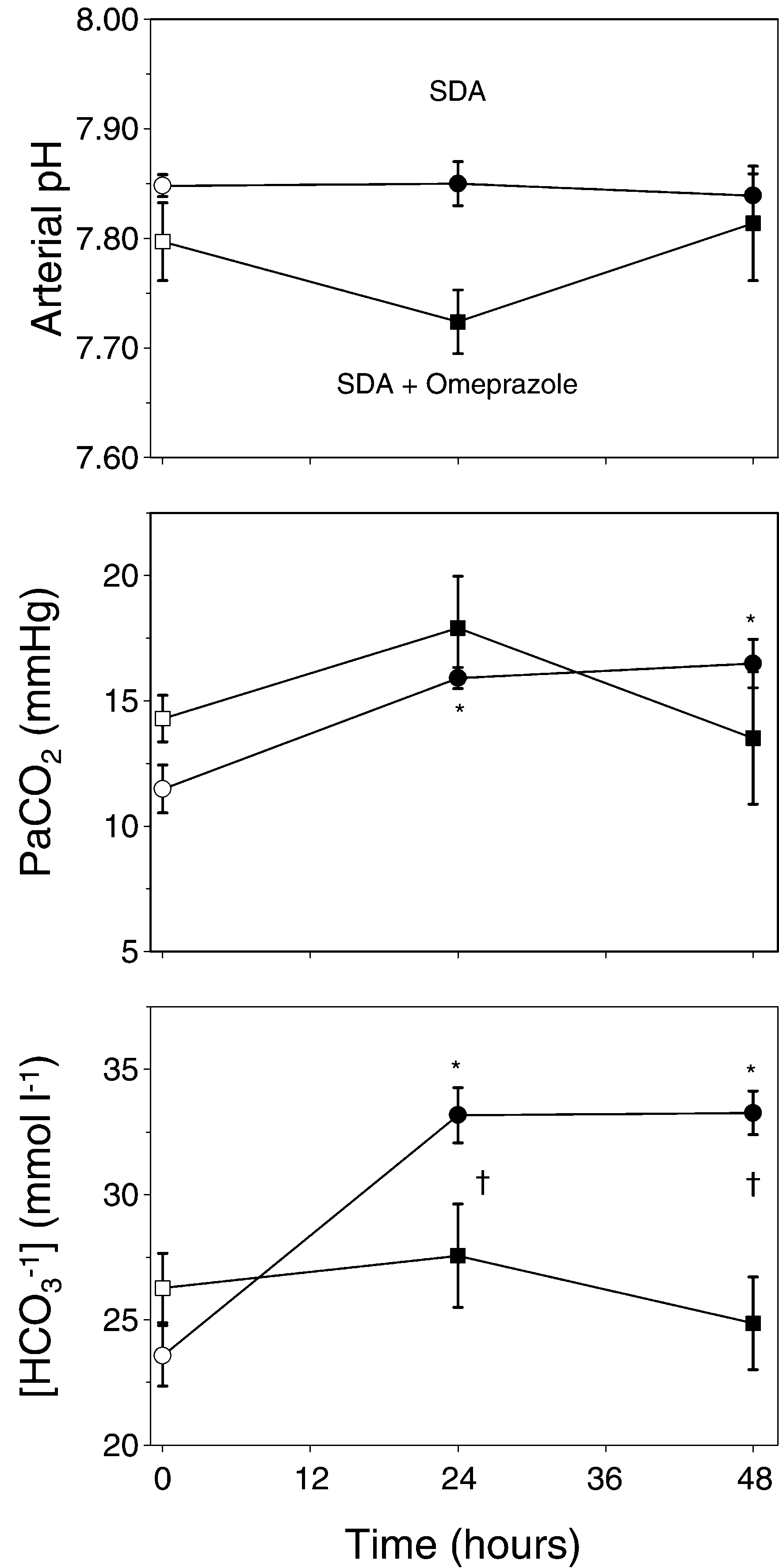 J.B. Andersen et al. / Comparative Biochemistry and Physiology Part A 135 (2003) 425–433
was associated with a substantial increase in pHafrom 7.74"0.03 to 7.91"0.02, at unchangedPaCO .
J.B. Andersen et al. / Comparative Biochemistry and Physiology Part A 135 (2003) 425–433
was associated with a substantial increase in pHafrom 7.74"0.03 to 7.91"0.02, at unchangedPaCO .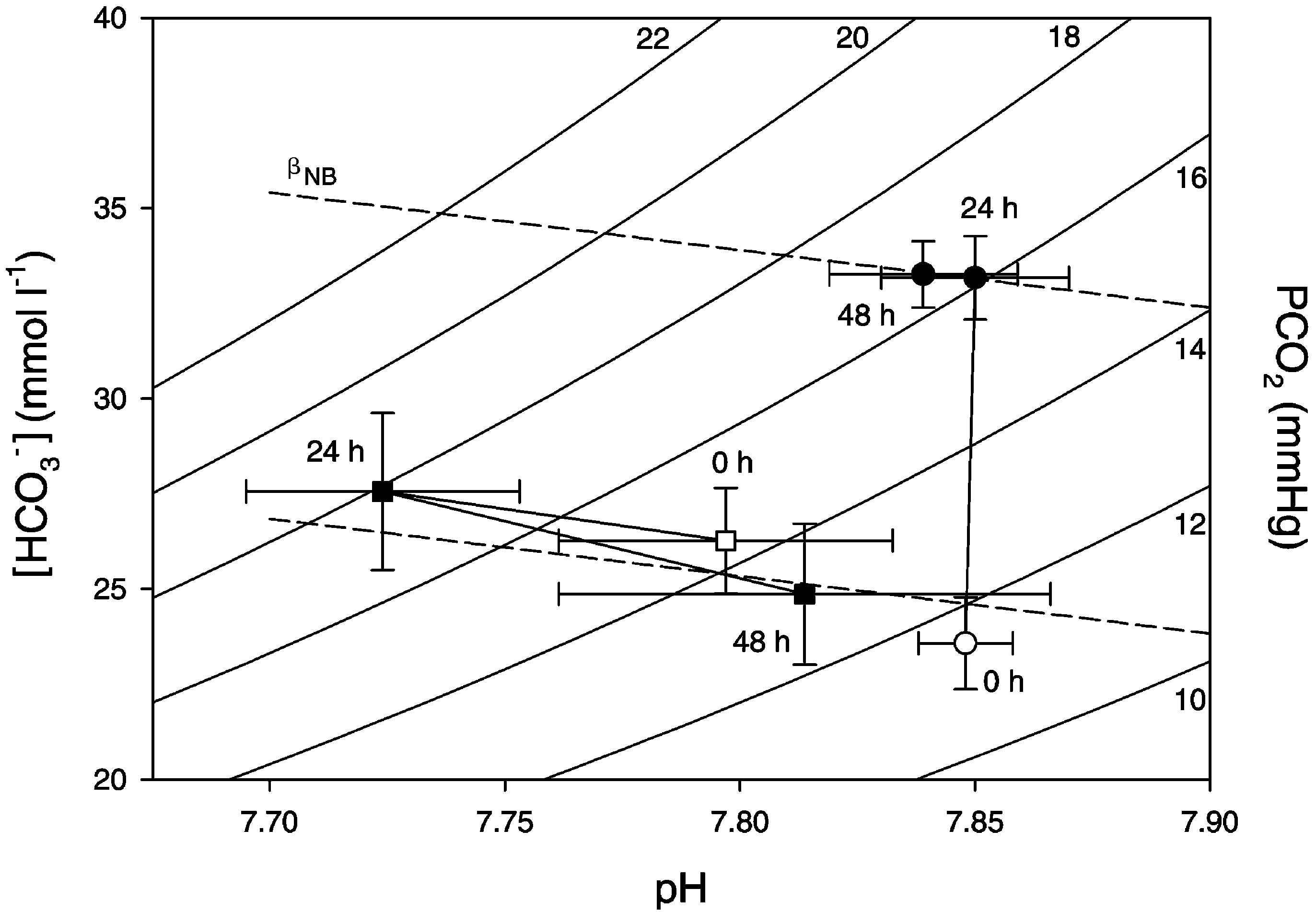 J.B. Andersen et al. / Comparative Biochemistry and Physiology Part A 135 (2003) 425–433
Fig. 2. Davenport diagram showing plasma w
and arterial pH during fasting and digestion in the toad Bufo marinus. Animals
treated with omeprazole are shown with the squares and untreated control animals are shown by circles. The Davenport diagram includestwo in vitro non-bicarbonate buffer lines (dotted lines, bNB) determined by and isobars for the partial pressureof CO2 in arterial blood (PaCO2, curved lines). Data are presented as mean "1 S.E.M. (Ns6 in each group).
J.B. Andersen et al. / Comparative Biochemistry and Physiology Part A 135 (2003) 425–433
Fig. 2. Davenport diagram showing plasma w
and arterial pH during fasting and digestion in the toad Bufo marinus. Animals
treated with omeprazole are shown with the squares and untreated control animals are shown by circles. The Davenport diagram includestwo in vitro non-bicarbonate buffer lines (dotted lines, bNB) determined by and isobars for the partial pressureof CO2 in arterial blood (PaCO2, curved lines). Data are presented as mean "1 S.E.M. (Ns6 in each group).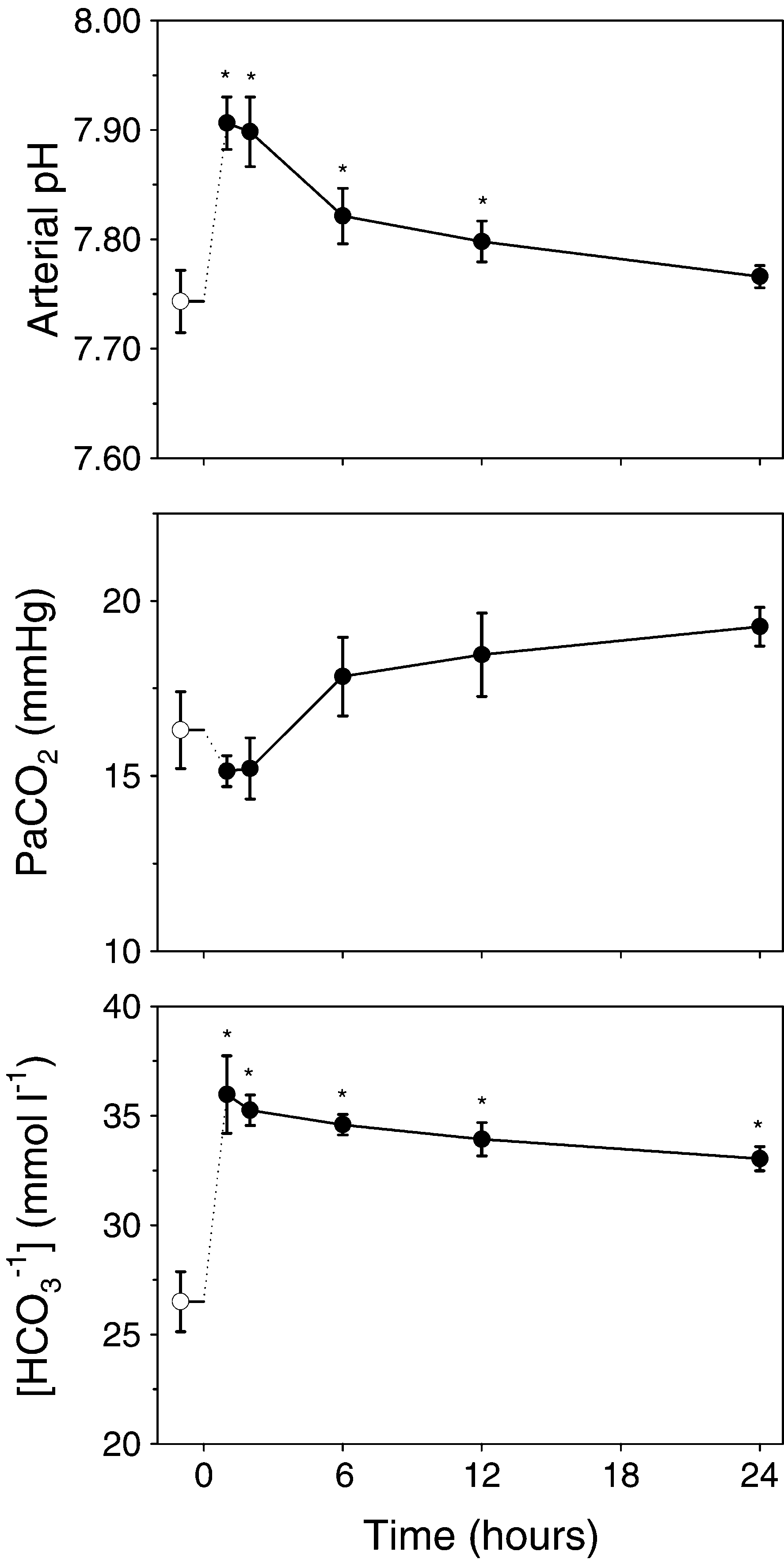 J.B. Andersen et al. / Comparative Biochemistry and Physiology Part A 135 (2003) 425–433
remains uncertain whether acid–base status offasting animals is affected by omeprazole. Inmammals, omeprazole is considered to be veryspecific and without side effects (e.g. and arterial acid–base parameters of fastingrats are not affected by omeprazole (Wang, Norlenand Haakanson, unpublished). Almost half of theomeprazole-treated toads vomited within 48 h afterforce feeding, and while their blood gas composi-tion did not differ from those completing digestion(data not shown), we excluded these animals fromthe study. It is likely that inhibition of gastric acidsecretion impaired the digestive ability and stimu-lated the emetic reflex, and secondary adverseeffects of omeprazole cannot be ruled out.
J.B. Andersen et al. / Comparative Biochemistry and Physiology Part A 135 (2003) 425–433
remains uncertain whether acid–base status offasting animals is affected by omeprazole. Inmammals, omeprazole is considered to be veryspecific and without side effects (e.g. and arterial acid–base parameters of fastingrats are not affected by omeprazole (Wang, Norlenand Haakanson, unpublished). Almost half of theomeprazole-treated toads vomited within 48 h afterforce feeding, and while their blood gas composi-tion did not differ from those completing digestion(data not shown), we excluded these animals fromthe study. It is likely that inhibition of gastric acidsecretion impaired the digestive ability and stimu-lated the emetic reflex, and secondary adverseeffects of omeprazole cannot be ruled out.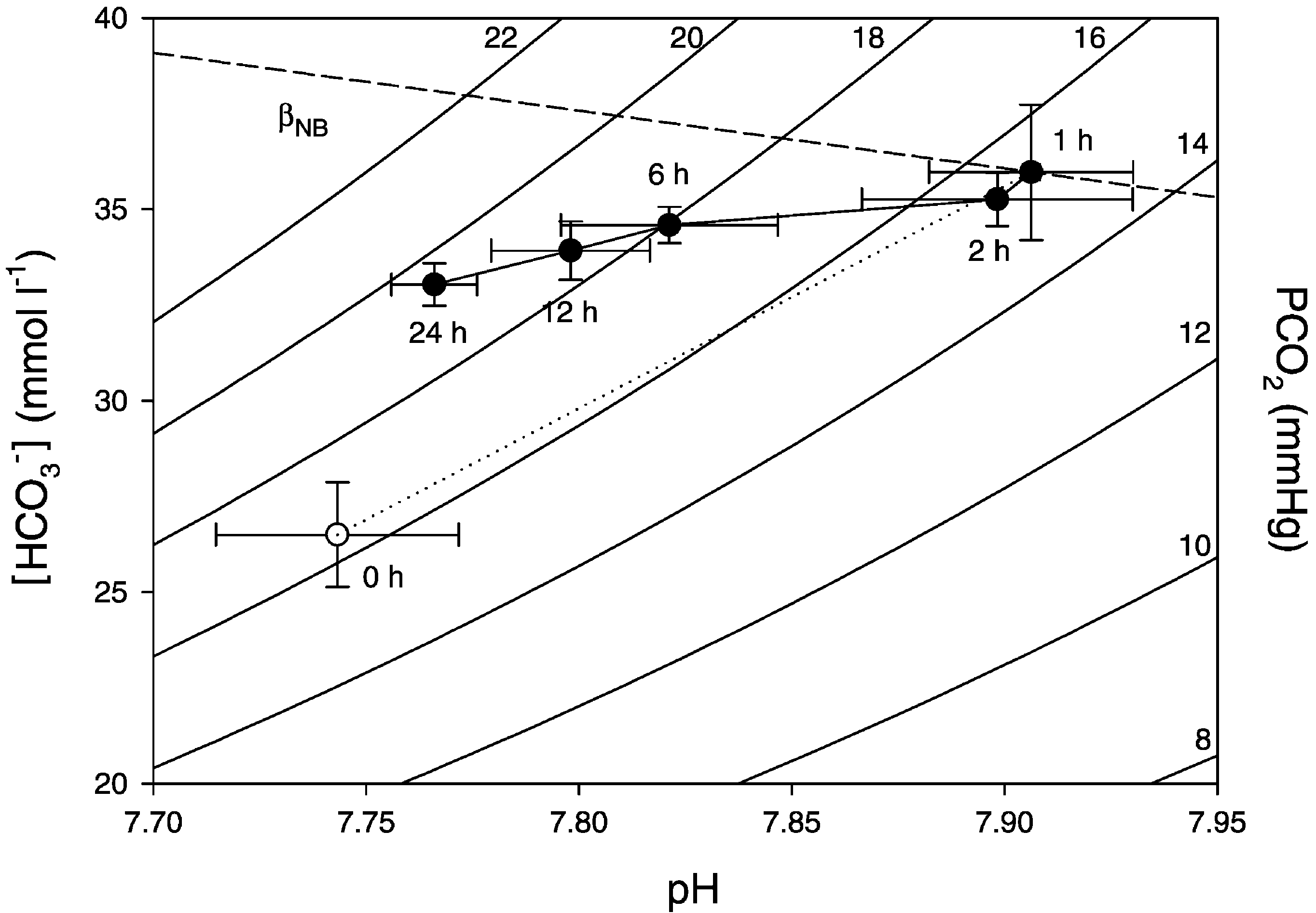 J.B. Andersen et al. / Comparative Biochemistry and Physiology Part A 135 (2003) 425–433
Fig. 4. Davenport diagram showing plasma w
and arterial pH of fasting Bufo marinus before (open symbols) and after (closed
symbols) a vascular injection of bicarbonate (6.9"0.04 mmol kgy1). The Davenport diagram includes two in vitro non-bicarbonatebuffer lines (dotted lines, bNB) determined by and isobars for the partial pressure of CO2 in arterial blood(PaCO2, curved lines). Data are presented as mean "1 S.E.M. (Ns6).
J.B. Andersen et al. / Comparative Biochemistry and Physiology Part A 135 (2003) 425–433
Fig. 4. Davenport diagram showing plasma w
and arterial pH of fasting Bufo marinus before (open symbols) and after (closed
symbols) a vascular injection of bicarbonate (6.9"0.04 mmol kgy1). The Davenport diagram includes two in vitro non-bicarbonatebuffer lines (dotted lines, bNB) determined by and isobars for the partial pressure of CO2 in arterial blood(PaCO2, curved lines). Data are presented as mean "1 S.E.M. (Ns6).