Do you want to buy antibiotics online without prescription? https://buyantibiotics24h.net/ - This is pharmacy online for you!
Fitoterapicos.com.ar
Acta Farm. Bonaerense 24 (4): 527-32 (2005)
Recibido el 11 de junio de 2005Aceptado el 19 de julio de 2005
Actividad Antiplasmódica In Vitro e Inhibición de la Formación
de la β-Hematina de Plantas Colombianas de la Familia Annonaceae.
Edison OSORIO 1, Gabriel ARANGO 1*, Edison GARCÍA 1, Katalina MUÑOZ 1, Grace RUIZ 2,
David GUTIÉRREZ 2, Marco Antonio PACO 2 & Alberto GIMÉNEZ 2
1 Grupo de Investigación en Sustancias Bioactivas (GISB). Sede de Investigación Universitaria SIU.Universidad de Antioquia. Calle 62 No. 52-59, Torre II, Lab 229. Medellín-Colombia.
2 Instituto de Investigaciones Fármaco Bioquímicas, Facultad de Ciencias Farmacéuticas y Bioquímicas,Universidad Mayor de San Andrés, Av. Saavedra 2224, La Paz-Bolivia.RESUMEN. Se evaluó la actividad antiplasmódica in vitro de 36 extractos provenientes de especies de la familia Annonaceae sobre las cepas de Plasmodium falciparum F32 sensible y W2 resistente a la cloroqui- na. Igualmente fue evaluada la capacidad de inhibición de la formación de la β-hematina (Ifβ-h) compa- rando ambas actividades por medio de un estudio de correlación estadístico. Cuatro extractos presentaron una potente actividad contra la cepa F32, y solamente tres mostraron actividad contra la cepa W2, siendo el extracto de hexano de tallos de Rollinia exsucca el mas activo en el estudio con una CI de 3.0 y 4.8 µg/ml sobre las cepas F32 y W2, respectivamente. Solamente el extracto de acetato de etilo de hojas de Desmopsis panamensis mostró actividad Ifβ-h e inhibición del crecimiento de ambas cepas del parásito en cultivo, mientras que el extracto de acetato de etilo de tallos de Rollinia pittieri presento actividad Ifβ-h e inhibición del crecimiento de la cepa F32. Fue observada una baja correlación entre ambas actividades. SUMMARY. “In Vitro Antiplasmodial Activity and Inhibition of β-Hematin Formation of Colombian Plants of theFamily Annonaceae”. The antiplasmodial activity of 36 plant extracts related to the Annonaceae family was test-ed on chloroquine sensitive strain F32 and chloroquine resistant strain W2 of Plasmodium falciparum. The capaci-ty of inhibition of β-hematin formation (Ifβ-h) was also evaluated comparing both activities by means of a statisti-cal correlation study. Four extracts presented a potent activity against F32 and three only showed activity againstW2, being the hexane stem bark extract of Rollinia exsucca the most active with CI50 values of 3.0 and 4.8 µg/mlagainst F32 and W2, respectively. Only the ethyl acetate leaves extract of Desmopsis panamensis showed Ifß-hactivity and growth inhibition of both strain, while the ethyl acetate stem bark extract of Rollinia pittieri presentsIfβ-h activity and growth inhibition of F32. A low correlation was observed among both activities. INTRODUCCIÓN
partir de especies del género Cinchona (Rubia-
La malaria es una enfermedad producida por
ceae) nativo de Sudamérica y otros derivados
parásitos del género Plasmodium. Según cálcu-
sintéticos desarrollados posteriormente que han
los de la Organización Mundial de la Salud
mostrado ser más efectivos, menos tóxicos y de
(OMS), anualmente ocurren entre 300 a 500 mi-
bajo costo 2. El desarrollo de resistencia de P.
llones de casos clínicos de la enfermedad, de
falciparum a los medicamentos disponibles for-
los cuales 1,5 a 2,7 millones son mortales, ade-
zó la investigación de nuevos compuestos con
más 2.400 millones de personas viven en regio-
actividad antiplasmódica y permitió el descubri-
nes de alto riesgo para su transmisión, lo que
miento de la mefloquina y la cloroquina, esta
hace de esta enfermedad la principal causa de
última aún se utiliza en algunas regiones de
morbilidad y mortalidad en 90 países ubicados
África y Sudamérica. El descubrimiento reciente
en las regiones tropicales y subtropicales del
más importante es la artemisinina, una sesqui-
mundo, especialmente en la región al sur del
terpenlactona con un puente endoperóxido ob-
Sahara en África, el sudeste de Asia y Latinoa-
tenida de Artemisia annua (Asteraceae) y el de-
sarrollo de sus derivados, el artemether y el ar-
El tratamiento de la malaria se ha realizado
tesunato, los cuales son medicamentos de rápi-
con diversos medicamentos que actúan sobre
da acción y efectivos contra cepas de P. falcipa-
estadios eritrocíticos del parásito, entre los que
rum resistentes a múltiples medicamentos 3. Sin
se encuentran la quinina, un alcaloide aislado a
embargo, la actividad de la artemisinina y sus
PALABRAS CLAVE: Actividad antiplasmódica, Annonaceae, β-hematina. KEY WORDS: Annonaceae, Antiplasmodial activity, β-hematin.
Autor a quien se debe de dirigir la correspondencia: E-mail: gjarango@quimbaya.udea.edu.co.
OSORIO E., ARANGO G., GARCÍA E., MUÑOZ K., RUIZ G., GUTIÉRREZ D., PACO M.A., GIMÉNEZ A.
derivados se ve afectada por su baja solubilidad
partir de plantas colombianas, fueron evaluados
y pobre biodisponibilidad. Además, aunque los
36 extractos provenientes de 6 especies de la fa-
derivados presentan una mejor absorción, rápi-
milia Annonaceae contra parásitos de P. falcipa-
da acción y mayor efectividad contra P. falcipa-rum resistentes y sensibles a cloroquina, igual-
rum multirresistente, presentan citotóxicidad y
mente, se evaluó la capacidad de inhibición de
la formación de la β-hematina, sustancia sintéti-
No obstante, el tratamiento de la malaria
ca idéntica a la hemozoina (pigmento malárico).
continúa como uno de los mayores retos para
Una de las hipótesis mayormente aceptada es
los programas de control, debido al fenómeno
que la cloroquina y otros antimaláricos quinolí-
de resistencia del parásito contra los medica-
nicos actúan inhibiendo la formación de la he-
mentos. Esta resistencia se debe a la capacidad
mozoina, por lo tanto este proceso permanece
del parásito para mutar genes específicos 6, a la
como un blanco atractivo para la búsqueda de
alta frecuencia de recombinación génica que da
nuevos compuestos antimaláricos 22. Se muestra
origen a poblaciones de parásitos con nuevos
estadísticamente la correlación entre ambas acti-
determinantes antigénicos y con modificaciones
en los sitios blanco para la acción de medica-mentos 7, a los sistemas de transporte activo es-
MATERIALES Y MÉTODOS
pecíficos para compuestos antimaláricos y a las
Procedimientos experimentales generales
prácticas clínicas inadecuadas como el uso de
El ensayo de inhibición de la formación de
antimaláricos profilácticos, tratamientos incon-
la β-hematina fue desarrollado por espectrofoto-
clusos o con dosis sub-terapéuticas 8. La meflo-
metría ultravioleta-visible utilizando un equipo
quina se utilizó por más de 10 años, pero se re-
Spectronic® Genesys 2 y una centrífuga 5415
gistró resistencia en el sudeste de Asia con resis-
Eppendorf, Brinkmann. Los espectros IR se rea-
tencia cruzada a quinina y con efectos tóxicos
lizaron en un equipo Perkin-Elmer (FT-IR) utili-
considerables. Igualmente durante muchos años
zando un rango de barrido entre 4000 y 600
la cloroquina fue el tratamiento de elección para
cm–1. El pH se determino con un pHmetro mar-
la malaria, pero en 1957 se reportaron los pri-
meros casos de resistencia en Sudamérica y des-de entonces el fenómeno se expandió a toda
Químicos
América, África y Asia. En algunas regiones se
Los solventes utilizados para la elaboración
ha informado de resistencia a todos los antima-
de los extractos son de grado reactivo suministra-
láricos disponibles con excepción de los deriva-
dos por J.T. Baker. La hemina (C34H32ClFeN4O4)
dos de la artemisinina, los cuales aún se reser-
y el medicamento control difosfato de cloroquina
van para los casos de malaria que no responden
(CQ) fueron adquiridos en Sigma Chemical Co,
a los demás medicamentos 8. Esta resistencia a
St Louis, Mo. Los reactivos hidróxido de sodio,
los medicamentos actualmente disponibles ha
ácido acético, acetato de sodio trihidratado, di-
llevado a la necesidad de desarrollar nuevos
metilsulfóxido (DMSO) fueron obtenidos de Sig-
compuestos antimaláricos que permitan mejorar
El estudio sobre la medicina tradicional co-
Material vegetal y preparación de
mo fuente que conduce al descubrimiento de
extractos
nuevos agentes antiparasitarios, ha encontrado
Las especies de la familia Annonaceae Anno-
que plantas de la familia Annonaceae han sido
na muricata, Desmopsis panamensis, Pseudo-
utilizadas por comunidades colombianas como
malmea boyacana, Rollinia exsucca, Rollinia
antiparasitarias 9,10. Algunas especies de esta
pittieri y Xylopia aromática fueron recolectadas
gran familia, la cual comprende alrededor de
en el corregimiento de Lomas Aisladas del Mu-
120 géneros y más de 2000 especies, presentan
nicipio de Turbo (Antioquia, Colombia) por el
interesantes metabolitos con actividad biológica:
biólogo Fernando Alzate e identificado en el
polifenoles, aceites esenciales, terpenos, com-
Herbario de La Universidad de Antioquia (Vou-
puestos aromáticos 11, siendo particularmente
chers en Tabla 1). El material vegetal (tallos y
activos las acetogeninas 12, moléculas con am-
hojas) fue secado a 40 °C en estufa con circula-
plio espectro de acción anticancerígena 13, anti-
ción de aire, pulverizado y extraído exhaustiva-
parasitaria 14,15 e insecticida 16 y los alcaloides
mente por percolación con solventes orgánicos
de tipo bisbencilisoquinoleicos 17,18, protoberbe-
de diferente polaridad (hexano, acetato de etilo
rinas, oxoaporfínicos 19,20 y aporfínicos 21. En la
y metanol) hasta agotar el material, el solvente
investigación de nuevos agentes antimaláricos a
fue removido bajo presión reducida. Para el en-
acta farmacéutica bonaerense - vol. 24 n° 4 - año 2005
sayo de inhibición de la formación de la β-he-
medio RPMI 1640 suplementada con suero al
matina, los extractos en metanol fueron disueltos
10% y un hematocrito del 4% (Grupo sanguíneo
en una solución agua-etanol (30:70) y los de ex-
0, Rh+) a 37 °C en un medio anaeróbico. Los
tractos en hexano y acetato de etilo en DMSO.
extractos fueron disueltos en DMSO y la cloro-quina en agua para luego ser diluidos con el
Actividad antiplasmódica in vitro
mismo medio obteniéndose las concentraciones
La actividad antiplasmódica se realizó por el
requeridas (0.10 ; 1.0 y 10.0 µg/ml). Los cultivos
método de cultivo continuo in vitro desarrollado
fueron sincronizados con una parasitemia y un
en 1976 por Trager & Jensen 23. De acuerdo con
hematocrito del 1 y 2% respectivamente, estos
esta técnica las formas parasitarias de P. falcipa-
fueron alicuotados en un volumen de 100 µl en
rum cepa F32 sensible a la cloroquina y W2 re-
placas de 96 pozos por duplicado, además de
sistente a la cloroquina, fueron cultivadas en
100 µl de los extractos, y finalmente fueron in-
P. falciparum a Solvente Ifβ-h (%) d científico F32 b W2 c Tabla 1. Actividad antiplasmódica in vitro sobre P. falciparum (IC50) y porcentaje de inhibición de la formación de β-hematina (promedio ± SD) de extractos de especies de la familia Annonaceae. a Actividad antiplasmódica, % de Inhibición a 10 µg/ml (CI50, µg/ml ). b F32, cepa de P. falciparum sensible a CQ. c W2, cepa de P. falci- parum resistente a CQ. d Ifβ-h, porcentaje de inhibición de la formación de β-hematina.
OSORIO E., ARANGO G., GARCÍA E., MUÑOZ K., RUIZ G., GUTIÉRREZ D., PACO M.A., GIMÉNEZ A.
cubados a 37 °C por 48 horas. Pasado este tiem-
(CQ) fue utilizado como control positivo y su
po de incubación, se eliminó completamente la
actividad inhibitoria es expresada en términos
fase superior del cultivo, para realizar un frotis
de CI50, es decir la concentración de CQ nece-
del sedimento de cada alveolo, fijando luego
saria para la inhibición del 50% de la formación
con metanol y realizando la tinción con Giemsa,
de β-hematina, y es calculada mediante el pa-
estas placas fueron observadas en el microsco-
quete estadístico GraphPad Prism® demo, Ver-
pio, con lente de inmersión 100x, contando gló-
sión 4.00 para Windows, (GraphPad software,
bulos rojo no infectados (GRL) y glóbulos rojos
Inc, San Diego CA 2003). Los ensayos fueron re-
infectados (GRI), para obtener el % de Inhibi-
ción calculado mediante la fórmula [1]:
RESULTADOS Y DISCUSIÓN
Los resultados obtenidos en el estudio de ac-
tividad antiplasmódica in vitro de extractos deespecies de la familia Annonaceae son mostrados
El cálculo para hallar la Concentración Inhi-
en la Tabla 1. Los extractos fueron evaluados a
bitoria del 50% en la maduración de los esqui-
una concentración de 10 µg/ml y aquellos que
zontes (CI50), se realizó por un método gráfico
presentaron una inhibición mayor del 50% en el
mediante el programa Cricket Graph 1.3, consi-
crecimiento de las cepas de P. falciparum sensi-
derándose como activos aquellos que presenta-
ble (F32) y resistente (W2) a CQ, se les calculó la
CI50 y fueron considerados como extractos depotente actividad antiplasmódica. Los extractos
Inhibición de la formación de la β-hematina
de hojas (EtOAc) de D. panamensis, tallos (EtO-
Para el ensayo de inhibición de la formación
Ac) de P. boyacana, tallos (Hexano) de R. exsuc-
de la β-hematina se utilizó el método de Bael-
ca y tallos (EtOAc) de R. pittieri, mostraron acti-
mans et. al. 24 con algunas modificaciones. En
vidad contra la cepa F32, y solamente los extrac-
resumen, la síntesis de la β-hematina fue realiza-
tos de hojas (Hexano y EtOAc) de D. panamen-
da con una mezcla de 100 µl de hemina 6,5 mM
sis y tallos (Hexano) de R. exsucca mostraron ac-
recién preparada disuelta en una solución de
tividad en la cepa W2. Este último extracto resul-
NaOH 0,2 N, 50 µl de ácido acético glacial 17,4
tó ser el más activo en el estudio de actividad an-
M, 50 µl de H2O destilada y 200 µl de tampón
tiplasmódica con una CI50 de 3,0 y 4,8 µg/ml en
acetato de sodio trihidratado 3 M, pH final apro-
las cepas F32 y W2 de P. falciparum respectiva-
ximadamente 4,0, fue incuba por 1,h a 60 °C.
mente. La CI50 de la CQ usada como medica-
Posteriormente fue centrifugada a 12000 rpm
mento control en el ensayo fue de 30 nM contra
durante 10 min, luego de descartar el sobrena-
la cepa F32 y 51 nM contra la cepa W2.
dante, el precipitado es lavado 3 veces con 200
Igualmente se evaluó la actividad de inhibi-
µl de DMSO para remover la hemina no reac-
ción de la formación de la β-hematina (Ifβ-h) a
cionante. El sólido β-hematina obtenido fue di-
los diferentes extractos (Tabla 1). La relevancia
suelto en una solución de NaOH 0,1 N de la
de evaluar dicha actividad se basa en la siguien-
cual se toma una alícuota para la lectura espec-
te observación: Durante su ciclo intraeritrocítico,
trofotométrica a 386 nm y corresponde al 100%
el parásito de la malaria degrada una gran canti-
de β-hematina sintetizada. La formación de β-
dad de hemoglobina presente en el citoplasma
hematina se monitoreo por espectroscopía IR-
de la célula hospedera (entre el 60-80%) 25. Du-
TF. Para la evaluación de la actividad inhibitoria
rante el proceso de proteolisis de hemoglobina
de los extractos, los 50 µl de H2O fueron rem-
en su vacuola digestiva ácida es producido he-
plazados por una solución del extracto corres-
me (Ferriprotoporfirina IX), un compuesto po-
pondiente a una concentración final de 2,5
tencialmente tóxico para el parásito. El parásito
mg/ml y con la lectura espectrofotométrica fue
carece de heme oxigenasa, enzima que dispone
calculado el porcentaje de inhibición mediante
del compuesto en otras especies, entonces lo
detoxifica en parte por su incorporación en unasustancia cristalina, inerte e insoluble denomina-
da hemozoina (o pigmento malárico) 26,27 y el
resto por procesos de degradación peroxidati-
vos. Esta función especializada hace de la va-
Donde Abmuestra y Abcontrol son la absorban-
cuola digestiva un blanco atractivo para la bús-
cia de la β-hematina con y sin el uso de extrac-
queda de nuevos compuestos antimaláricos, un
tos, respectivamente. El difosfato de cloroquina
compuesto capaz de inhibir la formación de he-
acta farmacéutica bonaerense - vol. 24 n° 4 - año 2005
mozoina, podría ser potencialmente letal para el
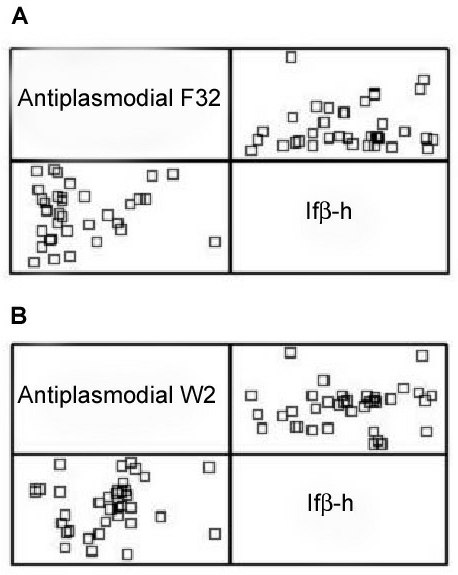
dows 4.1 (Statistical Graphics Corp, 1999) (Fig.
parásito. Para muchas sustancias, su habilidad
1). No hay una significante correlación entre las
de inhibir esta formación esta directamente rela-
actividades antiplasmódica sobre la cepa F32 y
cionada con su potencia antimalárica 28,29.
W2 con la Ifβ-h con un índice de correlación de
Una sustancia similar a la hemozoina, la β-
hematina, puede ser formada in vitro a partir de
La falta de correlación puede deberse a fac-
una solución de hemina (hidroxi-ferriprotoporfi-
tores relacionados con la incapacidad de los
rina IX) bajo ciertas condiciones de pH, tempe-
principios activos de los extractos de alcanzar el
ratura y concentración de sales que simulan el
sitio de formación de la hemozoína y de poder
ambiente de la vacuola digestiva 30. La β-hemati-
acumularse en la vacuola digestiva ácida del pa-
na sintética es espectrofotométrica y química-
rásito a concentraciones efectivas. Las moléculas
mente idéntica a la hemozoina, además conser-
deben de penetrar libremente a través de un
va las propiedades de solubilidad de la sustan-
complejo sistema de membranas: por parte del
cia nativa 31, siendo útil en el estudio y diseño
eritrocito, la membrana celular y la membrana
de nuevos agentes terapéuticos. La metodología
de la vacuola parasitófora y, por parte del pará-
aplicada para la formación de β-hematina se
sito, su membrana y la membrana de la vacuola
monitoreó por espectroscopía de IR-TF, en don-
digestiva 24. Así mismo, se ha demostrado la im-
de se distingue inequívocamente hemina de β-
portancia que tiene la acumulación de la droga
hematina 32. El espectro IR de β-hematina mos-
a nivel de la vacuola digestiva ácida del parási-
tró bandas a 1662 y 1209 cm–1 características de
to, en cuanto a la potencia de compuestos anti-
la unión hierro-carboxilato 27, las cuales no es-
maláricos y el efecto que ejerce esta acumula-
tán presentes en el espectro IR de la hemina
ción sobre la actividad de inhibición de la for-
mación de la hemozoína 33. Factores como el
Entre los 36 extractos examinados por su ac-
pH ácido de la vacuola digestiva y los sitios dis-
tividad Ifβ-h (Tabla 1), hojas (EtOAc) de A. mu-
ponibles en donde la droga tenga la posibilidad
ricata, hojas (EtOAc) de D. panamensis, hojas
de unirse al heme, pueden afectar dicha acumu-
(Hexano) de R. exsucca, hojas (Hexano) y tallos
lación. Por supuesto, otra posible alternativa
(EtOAc) de R. pittieri presentaron porcentajes de
puede deberse a la posibilidad de que los com-
inhibición mayores del 85% cuando fueron eva-
puestos activos no interfieran con la formación
luados a una concentración de 2,5 mg/ml. La
de la hemozoina y su mecanismo de acción sea
CI50 de la CQ fue de 1,15 mg/ml (2,24 mM) yestuvo en concordancia con previos reportes endonde se muestra que un exceso molar de CQsobre la hemina previene la formación de la β-hematina 24. Bajo las condiciones de la presenteinvestigación, (pH 4,0, 60 °C), 1,80 mg/ml (3,5mM) de CQ inhibió el 96% la formación de la β-hematina. Solamente el extracto de las hojas deD. panamensis mostró actividad Ifβ-h e inhibi-ción del crecimiento de ambas cepas del parási-to en cultivo, mientras que el extracto de tallosde R. pittieri presentó actividad Ifβ-h e inhibi-ción del crecimiento de la cepa F32 sensible aCQ. Para estos 2 últimos extractos, la correla-ción entre la actividad Ifβ-h y la actividad anti-plasmódica es clara, sin embargo, para los de-más extractos considerados como extractos depotente actividad antiplasmódica, la correlacióncon la actividad Ifβ-h no fue evidente. Estos ex-tractos mostraron porcentajes de inhibición me-nores del 65%, e inclusive extractos como el de
Figura 1. Correlación de la actividad antiplasmódica y
tallos de R. exsucca presentó una actividad Ifβ-h
la actividad de inhibición de la formación de β-hemati-
cercana a 23%. Para estudiar la relación entre
na. (A) Actividad antiplasmódica calculada sobre la ce-
estas dos actividades, se realizó un análisis de
pa F32, índice de correlación de 0.09, p-value = 0.60.
correlación mediante el método de Pearson uti-
(B) Actividad antiplasmódica calculada sobre la cepa
lizando el programa Statgraphics Plus para Win-
W2, índice de correlación de 0.11, p-value = 0.51.
OSORIO E., ARANGO G., GARCÍA E., MUÑOZ K., RUIZ G., GUTIÉRREZ D., PACO M.A., GIMÉNEZ A.
totalmente diferente. Al menos esto podría ser
tein & R. Antón (2001) J. Ethnopharmacol. 78:
el caso de extractos como el de tallos de R. ex-succa y hojas de Desmopsis panamensis, los
11. Leboeuf, M., A. Cave, P. Bhaumik, B. Mukher-
cuales mostraron potente actividad antiplasmó-
jee & R. Mukherjee (1982) Phytochemistry 21:
12. Cave, A., B. Figadere, A. Laurens & D. Cortes
CONCLUSIONES
(1997) “Acetogeninas from Annonaceae” (W. Herz, G.W. Kirbi, R.E. Moore & W. Steglich,
En anteriores reportes se había estudiado ya
la pertinencia de la inhibición de la formación
13. Oberlies, N.H., V.L., Croy, M.L., Jarrison, & J.L.
de β-hematina en relación a la detección de po-
McLaughlin (1997) Cancer Lett. 115: 73-9.
tenciales compuestos antimaláricos a partir de
14. Jaramillo, M.C., G.J. Arango, M.C. González,
extractos 24; sin embargo, se plantea ahora un
S.M. Robledo, I.D. Vélez (2000) Fitoterapia 71:
análisis estadístico que muestra una significativa
no correlación entre la actividad antiplasmódica
15. Février, A., M.E. Ferreira, A. Fournet, G. Yaluff,
in vitro y la inhibición de la formación de β-he-
A. Inchausti, A. Rojas de Arias, R. Hocquemiller
matina. A pesar de este resultado, la informa-
& A.I. Waechter (1999) Planta Med. 65: 47-9.
ción suministrada por el ensayo de actividad
16. Ahammadsahib, K.I., R.M. Hollingworth, J.P.
Ifβ-h es importante en la búsqueda de alternati-
McGovren, Y.H. Hui & J.L. McLaghlin (1993) Life Sci. 53: 1113-20.
vas terapéuticas contra la enfermedad de la ma-
17. Mambu, L., M.T. Martín, D. Razafimahefa, D.
laria, toda vez que exista la posibilidad de en-
Ramanitrashasimbola, P. Rasoanaivo & F. Frap-
contrar compuestos que interfieran con la for-
pier (2000) Planta Med. 66: 537-40.
mación de la hemozoina y que además, reúnan
18. Mahiou, V., F. Roblot, A. Fournet, & R. Hoc-
las características fisicoquímicas necesarias para
quemiller (2000) Phytochemistry 5 4: 709-16.
el transporte a través de membranas y de acu-
19. Ferreira, E., F. Roblot, A. Cavé, M.Q. Paulo &
A. Fournet (1996) J. Nat. Prod. 59: 438-40.
20. Del Rayo, M., G.C. Kirby, D.C. Warhurst, S.L. Agradecimientos. Este trabajo fue financiado por
Croft & J.D. Phillipson (2000) Planta Med. 66:
COLCIENCIAS (Contrato N° RC 108-2003) y CODI-
Universidad de Antioquia (Acta No. CPT 0313).
21. Nkunya, M.H., S.A. Jonker, J.J. Makangara, R.
Waibel & H. Achenbach (2000) PhytochemistryREFERENCIAS BIBLIOGRAFICAS 53: 1067-73.
1. WHO Expert Committee on Malaria (2000)
22. Egan, T.J. (2004) Drug Des. Rev. 1: 93-110. “Technical Report Series, Twentieth Report”.
23. Trager, W. & J.B. Jensen (1976) Natur. 263:
2. Marshall, E. (2000) Science 290: 437-9.
24. Baelmans, R., E. Deharo, G. Bourdy, V. Mu-
3. Meshnick, S. (1998) Med. Trop. 58: 13-7.
ñoz, C. Quenevo, M. Sauvain & H. Ginsburg
4. Galal, A., M. Ahmad, F. El-Feraly & A. McPhail
(2000) J. Ethnopharmacology 73: 271-5.
(1996) J. Nat. Prod. 59: 917-20.
25. Francis, S., D. Sullivan & D. Goldberg (1997)
5. Beekman, A., A. Barentsen & Woerdenbag
Annu. Rev. Microbiol. 51: 97-123.
(1997) J. Nat. Prod. 60: 325-30.
26. Slater, A., W. Swiggard, B. Orton, W.D. Flitter,
6. Marsh, K. (1999) “Genetic approaches to the
D.E. Goldberg, A. Cerami & G.B. Henderson
determinations of drugs pathogenesis and in-
(1991) Proc. Nat.l Acad. Sc.i USA 88: 325-9.
fectivity in Plasmodium falciparum malaria”,
27. Goldberg, D. & A. Slater (1992) Parasitol. To-
en: “Malaria molecular and clinical aspects”,
day. 8: 280-3.
28. Egan, T.J., D.C. Ross & P.A. Adams (1994)
FEBS Lett. 352: 54-7.
7. Walliker, D., H. Babiker & L. Cartwright (1998)
29. Egan, T.J. & H.M. Marques (1999) Coord.“Malaria: Parasite biology, pathogenesis, andChem. Rev. 192: 493-517. protection”, Ed. ASM Press, Washington, D.C.,
30. Basilico, N., E. Pagani, D. Monti, P. Olliaro &
D. Taramellia (1998) J. Antimicrob. Chemoth.
8. Phillips, R.S. (2001) Clin. Microbiol. Rev. 14: 42: 55-60.
31. Pagola, S., P. Stephens, D. Bohle, A. Kosar &
9. Blair, S., A. Correa, B. Madrigal, C. Zuluaga &
S. Madsen (2000) Nature 404: 307-10.
H. Franco (1991) “Plantas antimaláricas, una
32. Basilico, N., D. Monti, P. Olliaro & D. Tarame-
revisión bibliográfica”, Ed. Universidad de An-
lli (1997) FEBS Lett. 409: 297-9.
33. Hawley, S.R., P.G. Bray, M. Mungthin, J.D. At-
10. Weniger, B., S. Robledo, G. Arango, E. Deha-
kinson, P.M. O’Neill & S.A. Ward (1998) Anti-
ro, R. Aragon, V. Munoz, J. Callapa, A. Lobs-
microb Agents Chemother. 42: 682-6.
Cocoons and Nor’easters This article is from the “Edifying the Body” section of the Church of God BigSandy’s Web site, churchofgodbigsandy.com. It was posted for the weekend By Lenny Cacchio LEE’S SUMMIT, Mo.—Social commentators sometimes refer to a conceptknown as cocooning. According to trend forecaster Faith Popcorn (yes, that’sher real name), “Cocooning is about stayin
Thesis proposal CSC 2013 Rational antitubercular drug design for targeting the NADH/NAD cofactor bindingsite of enoyl-ACP of Mycobacterium tuberculosis Thesis supervisor: E-mail address: PhD School name: Physics, Chemistry & Material Sciences (SDM) Research Laboratory: Laboratoire de Chimie de Coordination du CNRS UPR 8241 Laboratory website: Scientific domain: Organic
 acta farmacéutica bonaerense - vol. 24 n° 4 - año 2005
mozoina, podría ser potencialmente letal para el
dows 4.1 (Statistical Graphics Corp, 1999) (Fig.
acta farmacéutica bonaerense - vol. 24 n° 4 - año 2005
mozoina, podría ser potencialmente letal para el
dows 4.1 (Statistical Graphics Corp, 1999) (Fig.