Do you want to buy antibiotics online without prescription? https://buyantibiotics24h.net/ - This is pharmacy online for you!
Doi:10.1016/s0300-9084(02)00021-4
Techniques for the removal of marker genes from transgenic plants
Charles P. Scutt a,*, Elena Zubko b, Peter Meyer b
a Reproduction et Développement des Plantes, École Normale Supérieure de Lyon, 46, allée d’Italie, 69364 Lyon cedex 07, Franceb Centre for Plant Sciences, University of Leeds, Leeds LS2 9JT, UK
Received 30 August 2002; accepted 24 October 2002
Abstract
The presence of marker genes encoding antibiotic or herbicide resistances in genetically modified plants poses a number of problems.
Various techniques are under development for the removal of unwanted marker genes, while leaving required transgenes in place. The aim ofthis brief review is to describe the principal methods used for marker gene removal, concentrating on the most recent and promisinginnovations in this technology.
2002 Éditions scientifiques et médicales Elsevier SAS and Société française de biochimie et biologie moléculaire. All rights reserved. Keywords: Marker-gene; Antibiotic; Herbicide; Genetically modified organism
1. Introduction
publicity related to the presence of unnecessary marker genesas sufficient reason to warrant their removal.
The addition of genes conferring desired traits to plants
In addition to environmental and health concerns, there
also requires the inclusion of marker genes that enable the
are also practical reasons for the removal of unnecessary
selection of transformed plant cells and tissues. These select-
marker genes from plants. Both in fundamental and applied
able markers are conditionally dominant genes that confer
research, there is frequently a need to add two or more
the ability to grow in the presence of applied selective agents
transgenes to the same plant line. One method for the serial
that are toxic to plant cells, or inhibitory to plant growth, such
transformation of plants involves the use of two or more
as antibiotics and herbicides. Following transformation, the
different selectable markers. However, the number of marker
continued presence of marker genes in genetically modified
genes available is limited and not all of these are well adapted
plants usually becomes unnecessary and may also be unde-
to all transformable plant species. The combination of sev-
sirable. Herbicide resistance marker genes in transgenic cropplants, for example, could escape to wild relatives of the crop
eral nuclear transgenes can also be achieved through sexual
through the transfer of pollen, potentially leading to the
crosses following the transformation of independent plant
spread of herbicide resistance in wild plant populations
lines. However, this is not possible in plants that must be
The presence of antibiotic resistance markers in transgenic
propagated by vegetative means. These include non-sexually
plants intended for human or animal consumption may also
reproducing plants and highly heterozygous varieties whose
be a cause for concern. Fears have been expressed that such
genetic backgrounds would be greatly changed by sexual
genes may be transferred horizontally to microorganisms of
reproduction. Examples of such vegetatively propagated
the gut flora of man or animals and lead to the spread of
plants include: apple, hybrid aspen, banana, cassava, euca-
antibiotic resistances in pathogenic microorganisms. Though
lyptus, grapevine, potato and strawberries In addition,
extensive studies have failed to detect a quantifiable risk of
the combination of transgenes by sexual crosses may be slow,
this occurrence many biotechnologists view the negative
particularly in trees species. The possibility of removingunwanted marker genes following plant transformation al-lows the same marker to be used for the sequential addition offurther transgenes. A second practical reason for the removal
* Corresponding author. Tel.: +33-4-72-72-86-03;
of marker genes relates to the greater possibility of instability
E-mail address: Charlie.Scutt@ens-lyon.fr (C. Scutt)
of transgene expression if several homologous marker gene
2002 Éditions scientifiques et médicales Elsevier SAS and Société française de biochimie et biologie moléculaire. All rights reserved. PII: S 0 3 0 0 - 9 0 8 4 ( 0 2 ) 0 0 0 2 1 - 4
C. Scutt et al. / Biochimie 84 (2002) 1119–1126
Table 1Marker genes and selective agents used for plant transformation. cp Can be used for chloroplast transformation
Serratia marcescens, KlebsiellaArabidopsis thaliana, Nicotiana tabacumE. coli TN5, Streptoalloteichus hindustanusKlebsiella pneumoniae var. iozaenaeAgrobacterium CP4, maize, Petunia
copies are present in the same plant Multiple copies of
plants, concentrating particularly on the more recent and
marker genes could potentially lead to the silencing of the
required transgenes through homology-dependent gene si-lencing mechanisms.
The problems associated with the presence of marker
2. Selectable marker genes used for plant
genes in transgenic plants have been known for quite some
transformation
time and various studies over the last decade have demon-strated methods for the removal of these, while leaving the
Approximately 25 marker genes, mostly conferring resis-
desired transgenes in place. All of the methods published up
tance to antibiotics or herbicides, have been successfully
until recently have suffered from various drawbacks limiting
used for plant transformation ). In addition, a num-
their efficiency or widespread applicability. Recently, how-
ber of so-called marker gene-free approaches to plant trans-
ever, a number of studies have presented methods that seem
formation have been developed Selection of transformed
to offer advantages over earlier techniques. These include
tissues in these systems is based on genes that confer the
methods for the removal of nuclear marker genes by intrach-
ability to proliferate or differentiate in the absence of some
romosomal recombination, or using inducible heterologous
otherwise essential factor, such as a necessary exogenous
recombinases, in addition to novel methods for the removal
plant hormone used in tissue culture. The gene that has so far
of chloroplast marker genes. The aim of this brief review is to
been most widely used in such an approach is the ipt gene
describe and compare the different techniques that have been
from the Ti plasmid of Agrobacterium tumefaciens, encoding
tested for the removal of marker genes from transgenic
the enzyme isopentyl transferase This enzyme cata-
C. Scutt et al. / Biochimie 84 (2002) 1119–1126
lyzes the synthesis of isopentyl AMP, a precursor of cytoki-
sexual crosses with a recombinase-expressing transformant
nins. The excessive level of cytokinins produced in plant
In either case, both the recombinase gene and its own
tissues constitutively expressing the ipt gene leads to a pro-
associated marker gene must subsequently be separated from
liferation of these tissues on hormone-free media. Plant tis-
the desired trait gene by genetic segregation. Two major
sues over-expressing the ipt gene exhibit an “extreme shooty
problems have been reported that limit the applications of
phenotype” characterized by a loss of apical dominance and
these simple recombinase systems. Firstly, all of these sys-
an inability to produce roots. The removal of the ipt gene can
tems require sexual crosses for the removal of recombinase
be accomplished using one of the forms of technology for
genes and so cannot be used with vegetatively propagated
marker gene removal discussed in this review. Recently, ipt
plants. Secondly, the expression of microbial recombinases
genes of plant origin that also produce elevated cytokinin
for prolonged periods in plant cells may result in unwanted
levels when over-expressed have been identified through
changes to the genome at sites removed from transgene
insertions. The use of microbial recombinases for marker
A further type of gene that can be specifically of use in
gene removal, however, continues in more refined systems
advanced strategies for marker gene elimination acts as a
such as the MAT and CLX vector systems discussed
dominant negative selective marker The proteins en-
below. A further advanced use of the Cre-lox recombination
coded by such genes act to inhibit the growth of plant tissues
system exploits a transformation cassette designed to elimi-
in the presence of appropriate selective agents. Under selec-
nate multiple tandem insertions of transgenes and to remove
tive conditions, these dominant negative markers may be
used to identify plant tissues that have lost their marker genesthrough a recombination event brought about by one of the
3.2. Transposable element-based systems
techniques discussed in this review. The tms2 gene of Agro-bacterium tumefaciens, e.g., has been used as such a domi-
Heterologous plant transposons have also been used for
nant negative marker This gene encodes the enzyme
the removal of marker genes In one such system, the
indolacetamide amidohydrolase that converts napthaline ac-
maize Ac transposable element was engineered to contain the
etemide (NAM) into the auxin NAA. Plants expressing tms2ipt gene, conferring a selectable “extreme shooty phenotype”
are unable to root on media containing NAM due to elevated
The Ac element encodes its own transposase and so its
excision conveniently removes this gene along with the iptmarker gene thereby obviating the need for sexualreproduction steps in the procedure. However, transposon-
3. The removal of marker genes from the plant nuclear
based systems of marker gene removal suffer from a number
of disadvantages. Their efficiency is low, partly due to thetendency of transposable elements to reinsert elsewhere in
3.1. Simple microbial recombinase-based systems
the genome. Excision of transposons is frequently imprecise,and repeated cycles of insertion and excision may lead to the
One of the earliest techniques tested for marker gene
generation of mutations at numerous unknown loci. The
removal involved the heterologous expression of microbial
continued presence of heterologous transposons may also
recombinase enzymes in plants to excise marker transgenes
lead to genomic instability in transgenic plants. For these
that were flanked by microbial recombination sequences.
reasons, transposon-based systems seem to be currently less
The general method employed for this is illustrated in
favored as a means of the removal of marker genes.
. For example, the Cre recombinase enzyme ofbacteriophage P1 has been used to excise marker genes
cloned between pairs of 34 bp directly repeated loxP recom-bination sites Such excision events are precise and leave
A further conceptually very simple method for marker
one loxP site in place. Other microbial recombinase enzymes
gene removal is based on the co-transformation of plants
that have been similarly used to remove marker genes from
using two distinct transgene constructs present in the same
transformed plants include the yeast FLP and R recombi-
transformed line of A. One of these con-
nases The FLP recombinase, encoded by a gene of the
structs contains the selectable marker transgene to be used,
Saccharomyces cerevisiae 2µ plasmid, catalyzes the recom-
while the other includes the desired trait transgene, itself
binatorial excision of sequences flanked by directly repeated
unlinked to any marker gene. In a variant of this technique,
FRT sites. The R recombinase of Zygosaccharomyces rouxii,
these two transgenes are inserted into two different T-DNA
acts similarly to catalyze recombinatorial excision between
elements present in the same “super-binary” plant transfor-
mation vector Co-transformation methods for marker
In early studies, the introduction of microbial recombi-
gene removal are based on the principle that a proportion of
nase genes into plant lines carrying desired trait genes was
transformed plants carrying the selectable marker gene will
achieved by re-transformation of these either with a recom-
also have integrated the required trait transgene at a second,
binase gene linked to a further selectable marker gene, or by
unlinked insertion site. Marker genes can subsequently be
C. Scutt et al. / Biochimie 84 (2002) 1119–1126
Fig. 1. Transgene constructions used for the removal of marker genes from transgenic plants. (A) The simple use of microbial recombinases such as Cre, FLPand R (B) An Ac Transposon-based method for the removal of nuclear marker genes (C) The intrachromosomal recombination method for nuclearmarker gene removal (D) The GST-MAT vector system (E) The CRX vector system (F) Removal of chloroplast marker genes by homologousrecombination Ac = maize Activator transposable element, CDS = coding sequence, GFP = gene coding Green Fluorescent Protein. GST promoter =glutathione S-tranferase promoter, ipt = isopentyl transferase gene, LB and RB = left and right Agrobacterium tumefaciens T-DNA border sequences, nptII =neomycin phosphotransferase II gene, TBS = transformation booster sequence, tms2 = indolacetamide amidohydrolase gene, XVE = estrogen-activated hybridtranscriptional regulator gene
removed from such plants by genetic segregation. Co-
Zubko et al. tested the efficiency of a pair of 352 bp attP
transformation methods suffer from the obvious inefficiency
regions from bacteriophage k as substrates for ICR in plants.
that only a proportion of plants carrying the selectable
During the integration of the k genome into the E. coli
marker will also carry the desired trait gene at an unlinked
chromosome, the phage k attP region recombines with a
site. Furthermore, as for the simple use of heterologous
bacterial attB site over a pair of homologous core sequences.
recombination systems, co-transformation methods cannot
The process of bacteriophage integration involves a phage-
be used for vegetatively propagated plants.
encoded k integrase and a bacterially encoded IntegrationHost Factor (IHF). The construction used for plant transfor-
3.4. An intrachromosomal recombination (ICR) system
mation in the studies of Zubko et al. containeda group of three marker and reporter genes flanked by a pair
A more recently devised alternative approach to the re-
of directly repeated attP sites. This entire element was situ-
moval of nuclear transgene markers exploits the natural
ated adjacent to a copy of the transformation booster se-
nuclear recombination systems present in plants Re-
quence (TBS) from Petunia hybrida and a test transgene
moval of marker genes by this approach is based on intrach-
conferring a desired trait. The TBS has been found to in-
romosomal recombination (ICR) between two directly re-
crease the frequency of both ICR and illegitimate recombi-
peated sequences flanking the marker gene to be excised.
nation events in Petunia, Nicotiana and maize In this
C. Scutt et al. / Biochimie 84 (2002) 1119–1126
study, transformed tobacco calli were initially selected on
However, in its present form, it does involve a two-stage
kanamycin-containing media and subsequently cultured on
procedure to select transgenic calli. Calli are transferred from
kanamycin-free media to allow for the loss of the nptII gene
selective to non-selective media for propagation and then
by ICR. The detection of ICR events was based on the
re-transferred to a selective shoot-inducing medium to detect
acquisition of sensitivity to kanamycin, and confirmed by the
(white) tissue that has lost the marker gene. Such lengthy
loss of a negative selection tms2 gene marker. Two identical
propagation may increase the risk of somaclonal mutations
excision events from 11 initial transformed callus cultures
In addition, it has been pointed out that the activity of
were recovered, in which a 5.9 kb region containing the three
attP sequences as recombination substrates has yet to be
marker genes and precisely one of the two attP sites had been
demonstrated in a large range of plant species, and the
lost by ICR. Some illegitimate recombination events in sister
mechanism by which the recombination of these sequences
occurs in plants is not yet fully understood though the
ICR events in plants have previously been found to be very
recent results of Siebert and Puchta may go some way
rare, with only 10 such events detectable in all of the cells of
a 6-week old tobacco plant In the studies of Zubko et al. however, the use of attP sequences and the TBS seems
to have greatly increased the frequency of ICR events, de-spite the absence of the enzymes and co-factors necessary for
The MAT (multiautotransformation) vector system repre-
the recombination of attP sites in the phage k system. The
sents a highly sophisticated approach for the removal of
structure of the attP site may partially explain its apparent
nuclear marker genes In this system, a chosen trait
recombination-stimulating activity, as sequences containing
transgene is placed adjacent to a multigenic element flanked
a high A + T base composition have been found to favor both
by RS recombination sites A copy of the selectable
ICR and illegitimate recombination events in plants
ipt gene from A. tumefaciens is inserted between these re-
However, a further possible explanation for these results has
combinase sites, together with the yeast R recombinase gene
recently emerged from a study of repair to double-stranded
and this entire assembly is situated within a T-DNA element
DNA breaks in plants. Such DNA breaks were previously
for the Agrobacterium-mediated transformation of plant tis-
known to be repaired by recombination events, though this
sues. The MAT vector system allows the removal of the R
was thought to occur predominantly by illegitimate, rather
recombinase gene along with the ipt gene. The system does
than by homologous recombination. Siebert and Puchta
not, therefore, require any sexual crosses for the removal of
however, devised a system capable of measuring the relative
marker or recombinase genes, and recombinase expression in
frequencies of repairs to double-stranded breaks by homolo-
plant tissues is limited to a minimal period of time, thereby
gous and illegitimate recombination mechanisms. In this
reducing the possibility of any unwanted recombination ef-
study, pairs of double-stranded DNA breaks were generated
fects. In an earlier version of the MAT vector, R recombinase
in a plant transgene insertion by the transient expression of a
activity was constitutively up-regulated by the action of the
rare-cutting restriction enzyme. In the transgene insertion
CaMV 35S promoter. This system was found to incur a risk
used, a pair of rare restriction sites to be cut was flanked by
of marker gene excision before the selection of transformed
partial sequences of the uidA (GUS) reporter gene, of which
plant tissues could take place. To circumvent this problem, a
the central portion formed a pair of direct repeats. The induc-
more recent version of the MAT vector allows for a
tion of pairs of double-stranded DNA breaks led to the loss of
delay in the excision of the ipt and R recombinase genes. This
a marker gene situated between the two rare restriction sites.
is made possible by the use of a chemically inducible glu-
In cases where these breaks were then repaired by homolo-
tathione S-transferase promoter from maize to drive R re-
gous recombination of the repeated GUS sequences, rather
combinase gene expression. Once the positive selection of
than by non-homologous end-joining, an active GUS gene
transformed plant tissues showing an “extreme shooty phe-
was reconstituted. This study found that double-stranded
notype” has occurred, the excisive recombination of RS sites,
breaks could be repaired either by homologous recombina-
leading to a loss of the recombinase and marker genes, is
tion or by non-homologus end-joining, and that both of these
induced by treatment with the herbicide antidote “Safener”.
events occurred at very high frequencies. The high incidence
This two-step procedure using MAT vectors has been suc-
of ICR events noted in the studies of Zubko et al.
cessfully demonstrated for tobacco and hybrid aspen trans-
therefore, might be explained by invoking the involvement of
formation, both of which are accomplished using organogen-
a double-stranded break repair mechanism whose activity
esis for plant regeneration. Plant species for which current
was in some way stimulated by the presence of attP se-
transformation techniques require regeneration of trans-
formed embryos from embryogenic cultures were thought to
The ICR method of marker gene removal has the advan-
be potentially not amenable to selection using the ipt gene
tage of relative simplicity as it does not require the expres-
However, it has recently been demonstrated that trans-
sion of a heterologous recombinase. In addition, this tech-
formed rice plants can be regenerated from embryogenic
nique does not require any sexual reproduction steps and
cultures by the use of the MAT vector system In this
could therefore be used for vegetatively propagated plants.
case, transformed embryos that had lost the ipt marker gene
C. Scutt et al. / Biochimie 84 (2002) 1119–1126
but retained the desired transgene were selected directly in a
up to 104 copies per cell This considerable amplifica-
one-step procedure without the occurrence of an “extreme
tion can give a very high level of transgene expression, which
may be useful for applications requiring high concentrationsof proteins. Examples of these include the engineering of
3.6. The CLX chemically inducible system
drought-resistance, or the production of pharmaceuticals inplanta by molecular farming. The chloroplast genome is
In a further highly sophisticated approach to nuclear
uniquely transmitted through the female germ line in many
marker gene removal, the Cre-lox recombination system has
crops, reducing the possibility of transgene escape via polli-
been engineered to be chemically inducible Antibiotic
nation into local wild populations of plants. Examples of
selection using the CLX vector system for plant transforma-
chloroplast transgenes used to date include: the Cry gene,
tion and marker gene removal is based on an nptII gene
encoding Bacillus thuringiensis (Bt) toxin to confer insect
driven by a constitutive promoter. This nptII gene is
resistance and the hST gene encoding human somatatro-
positioned adjacent to a Cre-recombinase gene driven by the
hybrid, chemically inducible OLexA-46 promoter, and a hy-
Transformation of chloroplasts is currently performed by
brid XVE gene encoding the binding protein necessary
biolistic methods. Following the integration of transgenes
for the induction of Cre gene transcription These
into the chloroplast genome, a heterogeneous population of
three transcription units, with the exception of the constitu-
plastids will exist in transformed tissues, and selection using
tive promoter driving XVE gene expression, are flanked by a
a marker gene is required to produce homoplasmic plants in
pair of directly repeated loxP sites. Background expression
which the modified plastid genome has completely replaced
of the Cre gene in Agrobacterium is avoided by the incorpo-
the unmodified one. Transformation cassettes used for chlo-
ration of a plant intron. Following the Agrobacterium-
roplast transformation contain sequences homologous to two
mediated transformation of Arabidopsis root tissues using
adjacent regions of the chloroplast genome to allow the
the CLX vector system and the selection of transformed
integration of the transgenes by homologous recombination.
tissues on kanamycin-containing media, Cre recombinase
Selectable marker genes and desired trait transgenes are
placed between these homologous recombination sequences.
b-estradiol. As a result of Cre recombinase activity, the XVE
The removal of marker genes from the chloroplast genome is
coding sequence and the Cre and nptII genes were lost by
particularly important as their very high copy numbers could
precise excisive recombination between the loxP sites. This
otherwise lead to high levels of unwanted marker gene prod-
excision led to the close juxtaposition of the promoter previ-
ucts. A further argument for the removal of chloroplast ge-
ously driving XVE expression, and a previously promoter-
netic markers relates to the conservation of activity that often
less Green Fluorescent Protein (GFP) coding sequence, with
exists between chloroplast and bacterial promoters. This
concomitant activation of GFP expression. As with the MAT
could increase the risk of the horizontal transfer of functional
vector system, the use of the CLX vector for transformation
marker genes from plants to bacteria. Fewer resistance genes
and marker gene removal exposes plants to recombinase
are available for chloroplast than for nuclear transformation,
activity for the minimum possible time period, and does so
with most of the published studies based on the use of the
after an adequate period of time has elapsed to permit trans-
aadA gene The paucity of available selection
formant selection. The CLX vector system benefits also from
methods for chloroplast transformation further increases the
a particularly tightly regulated system of chemical induction
value of technology that enables the recycling of marker
The procedure could be used for vegetatively propa-
genes for the serial re-modification of a single transgenic
gated species and may be particularly well adapted to crop
species requiring transformation by the regeneration of em-bryo cultures. 4.1. Homologous recombination systems
As the integration of foreign transgenes into the chloro-
4. The removal of marker genes from the chloroplast
plast genome takes place by homologous recombination, it
was entirely logical to test this native plant mechanism as ameans for the removal of marker genes from the chloroplast
The genetic modification of chloroplasts can represent an
The first demonstration of this technique was
attractive alternative to engineering of the plant nuclear ge-
performed on the unicellular green alga, Chlamydomonas
nome for some applications Unlike the nuclear transfor-
reinharIamtham and Day then demonstrated its
mation of higher plants, chloroplast transformation takes
applicability to higher plants using a construction of three
place almost invariably by homologous recombination, re-
marker genes which shared two identical promoter se-
sulting in precise and predictable genetic modifications. The
quences of 174 bp and three identical terminator sequences
plastid genome is present in multiple copies in each or-
of 418 bp. Several different recombinative excision events
ganelle, and these can multiply to large numbers, particularly
were detected between the similar sets of promoter or termi-
in leaf tissues, such that a chloroplast transgene can exist in
nator sequences in the series of transgenic plants analyzed in
C. Scutt et al. / Biochimie 84 (2002) 1119–1126
these studies. After the removal of antibiotic selection, these
already exist. It seems highly likely that continued work in
excision events accumulated to high frequency, leading to a
this area will soon remove the question of unwanted marker
homoplastic, marker-free state in approximately 25% of
genes from the debate concerning the public acceptability of
transgenic lines in the next generation. Homoplastic marker-
transgenic crop plants. The techniques for marker gene re-
free plants may be identified in this technique by PCR or by
moval under development will also facilitate the more pre-
cise and subtle engineering of the plant genome, with wide-spread applications in both fundamental research and
4.2. Cre-lox recombination-based systems
Two recent studies have demonstrated that the Cre-lox
system can also be used for the removal of plastid transgene
Acknowledgements
markers These systems function essentially as forthe removal of nuclear transgenes by Cre-lox recombination.
C.P.S. is funded as a researcher of the Centre National de
A Cre-recombinase gene is expressed from a plant transfor-
la Recherche Scientifique (CNRS). The laboratory of Repro-
mation cassette integrated into the nuclear genome, while an
duction et Développement des Plantes (RDP) is funded
N-terminal chloroplast-directing signal sequence routes the
jointly by the CNRS, the Institut National de la Recherche
Cre recombinase protein that is produced to the plastids.
Agronomique, the École Normale Supérieure de Lyon and
Plastid transgene constructions for use with these methods of
the Université Claude Bernard-Lyon.
marker gene removal contain selectable marker genesflanked by loxP recombination sites. In one study dif-ferent results were noted depending on whether the Cre-
References
recombinase gene was introduced into the nuclear genome ofa chloroplast transformed line by direct transformation or by
P.J. Dale, Spread of engineered genes to wild relatives?, Plant Physi-
sexual crossing to a Cre recombinase transformant. In cases
J. Gressel, Indiscriminate use of selectable markers – sowing wild
where Cre was introduced by re-transformation, excisive
oats, Trends in Biotechnology 10 (1992) 382–382.
homologous recombination events between similar genetic
H.A. Kuiper, G.A. Kleter, H. Noteborn, E.J. Kok, Assessment of the
elements present in the transgene construction, as noted in
food safety issues related to genetically modified foods, Plant Journal
the studies of Iamtham and Day were observed at an
efficiency approximately equivalent to that of Cre-mediated
H. Ebinuma, K. Sugita, E. Matsunaga, M. Yamakado, Selection ofmarker-free transgenic plants using the isopentenyl transferase gene,
excision events. However, excision events by homologous
Proceedings of the National Academy of Sciences of the United States
recombination were not observed when the Cre gene was
introduced though sexual crosses. It is not currently clear
M. Matzke, A. Matzke, Differential inactivation and methylation of a
whether Cre-lox-based systems represent a considerable in-
transgene in plants by two suppressor loci containing homologous
crease in efficiency over homologous recombination for the
sequences, Plant Molecular Biology 16 (1991) 821–830.
J.R. Zuo, Q.W. Niu, Y. Ikeda, N.H. Chua, Marker-free transformation:
removal of chloroplast marker genes. One disadvantage of
increasing transformation frequency by the use of regeneration-
the current Cre recombinase-mediated methods for chloro-
promoting genes, Current Opinion in Biotechnology 13 (2002)
plast transgene removal is that they require the sexual cross-
ing of transformed plant lines to remove the nuclear-encoded
T. Kunkel, Q.W. Niu, Y.S. Chan, N.H. Chua, Inducible isopentenyl
recombinase gene and its associated genetic marker and so
transferase as a high-efficiency marker for plant transformation,
cannot be used for vegetatively propagated plants.
Nature Biotechnology 17 (1999) 916–919.
E. Zubko, C.J. Adams, I. Machaekova, J. Malbeck, C. Scollan,P. Meyer, Activation tagging identifies a gene from Petunia hybridaresponsible for the production of active cytokinins in plants, Plant
5. Conclusions
P. Hare, N.H. Chua, Excision of selectable marker genes from trans-genic plants, Nature Biotechnology 20 (2002) 575–580.
The field of marker gene removal continues to produce
[10] E. Zubko, C. Scutt, P. Meyer, Intrachromosomal recombination
new innovations. For example, the possibilities of increasing
between attP regions as a tool to remove selectable marker genes from
the number of different heterologous recombinase systems
tobacco transgenes, Nature Biotechnology 18 (2000) 442–445.
available by molecular evolution approaches have been dis-
[11] E.C. Dale, D.W. Ow, Gene-transfer with subsequent removal of the
selection gene from the host genome, Proceedings of the National
cussed, and new marker gene and marker-free strategies are
Academy of Sciences of the United States of America 88 (1991)
under development Work in progress aims to devise
systems that are efficient, rapid, precise, applicable to many
[12] K. Sugita, T. Kasahara, E. Matsunaga, H. Ebinuma, A transformation
plant species and transformation methods, do not require
vector for the production of marker-free transgenic plants containing a
sexual reproduction steps, minimize the risk of introducing
single copy transgene at high frequency, Plant Journal 22 (2000)461–469.
unwanted genetic changes, and leave the genome in a stable
[13] J.R. Zuo, Q.W. Niu, S.G. Moller, N.H. Chua, Chemical-regulated,
condition. Though research continues, it is clear that several
site-specific DNA excision in transgenic plants, Nature Biotechnol-
viable methods for the removal of unwanted marker genes
C. Scutt et al. / Biochimie 84 (2002) 1119–1126
[14] V. Srivastava, O.D. Anderson, D.W. Ow, Single-copy transgenic
[24] S. Endo, K. Sugita, M. Sakai, H. Tanaka, H. Ebinuma, Single-step
wheat generated through the resolution of complex integration pat-
transformation for generating marker-free transgenic rice using the
terns, Proceedings of the National Academy of Sciences of the United
ipt-type MAT vector system, Plant Journal 30 (2002) 115–122.
States of America 96 (1999) 11117–11121.
[25] J.R. Zuo, Q.W. Niu, N.H. Chua, An estrogen receptor-based transac-
[15] J. Yoder, A. Goldsbrough, Tranformation systems for generating
tivator XVE mediates highly inducible gene expression in transgenic
marker-free transgenic plants, Biotechnology 12 (1994) 263–267.
plants, Plant Journal 24 (2000) 265–273.
[16] M. De Block, D. Debrouwer, Two T-DNAs co-transformed into Bras-
[26] H. Daniell, M.S. Khan, L. Allison, Milestones in chloroplast genetic
sica napus by a double Agrobacterium infection are mainly integrated
engineering: an environmentally friendly era in biotechnology, Trends
at the same locus, Theoretical and Applied Genetics 82 (1991)
[27] M. Kota, H. Daniell, S. Varma, S.F. Garczynski, F. Gould, W.J. Moar,
[17] T. Komari, Y. Hiei, Y. Saito, N. Murai, T. Kumashiro, Vectors carrying
Overexpression of the Bacillus thuringiensis (Bt) Cry2Aa2 protein in
two separate T-DNAs for co-transformation of higher plants mediated
chloroplasts confers resistance to plants against susceptible and
by Agrobacterium tumefaciens and segregation of transformants free
Bt-resistant insects, Proceedings of the National Academy of Sciences
from selection markers, Plant Journal 10 (1996) 165–174.
of the United States of America 96 (1999) 1840–1845.
[28] J.M. Staub, B. Garcia, J. Graves, P.T.J. Hajdukiewicz, P. Hunter,
[18] H. Galliano, A.E. Muller, J.M. Lucht, P. Meyer, The transformation
N. Nehra, V. Paradkar, M. Schlittler, J.A. Carroll, L. Spatola, D. Ward,
booster sequence from Petunia hybrida is a retrotransposon derivative
G.N. Ye, D.A. Russell, High-yield production of a human therapeutic
that binds to the nuclear scaffold, Molecular and General Genetics
protein in tobacco chloroplasts, Nature Biotechnology 18 (2000)
[19] H. Puchta, P. Swoboda, S. Gal, M. Blot, B. Hohn, Somatic intrachro-
[29] N. Fischer, O. Stampacchia, K. Redding, J.D. Rochaix, Selectable
mosomal homologous recombination events in populations of plant
marker recycling in the chloroplast, Molecular and General, Genetics
siblings, Plant Molecular Biology 28 (1995) 281–292.
[20] A.E. Muller, Y. Kamisugi, R. Gruneberg, I. Niedenhof, R.J. Horold,
[30] S. Iamtham, A. Day, Removal of antibiotic resistance genes from
P. Meyer, Palindromic sequences and A+T-rich DNA elements pro-
transgenic tobacco plastids, Nature Biotechnology 18 (2000)
mote illegitimate recombination in Nicotiana tabacum, Journal of
Molecular Biology 291 (1999) 29–46.
[31] S. Corneille, K. Lutz, Z. Svab, P. Maliga, Efficient elimination of
[21] R. Siebert, H. Puchta, Efficient repair of genomic double-strand
selectable marker genes from the plastid genome by the CRE-lox
breaks by homologous recombination between directly repeated
site-specific recombination system, Plant Journal 27 (2001) 171–178.
sequences in the plant genome, Plant Cell 14 (2002) 1121–1131.
[32] P.T.J. Hajdukiewicz, L. Gilbertson, J.M. Staub, Multiple pathways for
[22] D.W. Ow, The right chemistry for marker gene removal?, Nature
Cre/lox-mediated recombination in plastids, Plant Journal 27 (2001)
[23] H. Ebinuma, K. Sugita, E. Matsunaga, M. Yamakado, A. Komamine,
[33] J.I. Yoder, A.P. Goldsbrough, Transformation systems for generating
Principle of MAT vector, Plant Biotechnology 14 (1997) 133–139.
marker-free transgenic plants, Biotechnology 12 (1994) 263–267.
HCTZ, HydroDIURIL®, Oretic®, and others), metolazone (Mykrox® and Zaroxolyn®). No known contraindications for Phytase. WARNINGS NDC 54799-917-05 Women who are pregnant, nursing or may become pregnant should consult their physician before using this product. Brief Summary of Prescribing Information PRECAUTIONS DESCRIPTION Zinc supplements may cause mild nausea in susc
APPLICATION FORM STUDENT INFORMATION (Please write in capital letters!) First Name(s) (please underline name used) GUARDIAN(S) INFORMATION Guardian 1 Guardian 2 Please tick if you are the child’s only guardian Square d'Argenteuil 5, B-1410 Waterloo, Belgium STUDENT INFORMATION Has the student had any type of learning support p


 Techniques for the removal of marker genes from transgenic plants
Charles P. Scutt a,*, Elena Zubko b, Peter Meyer b
a Reproduction et Développement des Plantes, École Normale Supérieure de Lyon, 46, allée d’Italie, 69364 Lyon cedex 07, France
b Centre for Plant Sciences, University of Leeds, Leeds LS2 9JT, UK
Received 30 August 2002; accepted 24 October 2002
Abstract
Techniques for the removal of marker genes from transgenic plants
Charles P. Scutt a,*, Elena Zubko b, Peter Meyer b
a Reproduction et Développement des Plantes, École Normale Supérieure de Lyon, 46, allée d’Italie, 69364 Lyon cedex 07, France
b Centre for Plant Sciences, University of Leeds, Leeds LS2 9JT, UK
Received 30 August 2002; accepted 24 October 2002
Abstract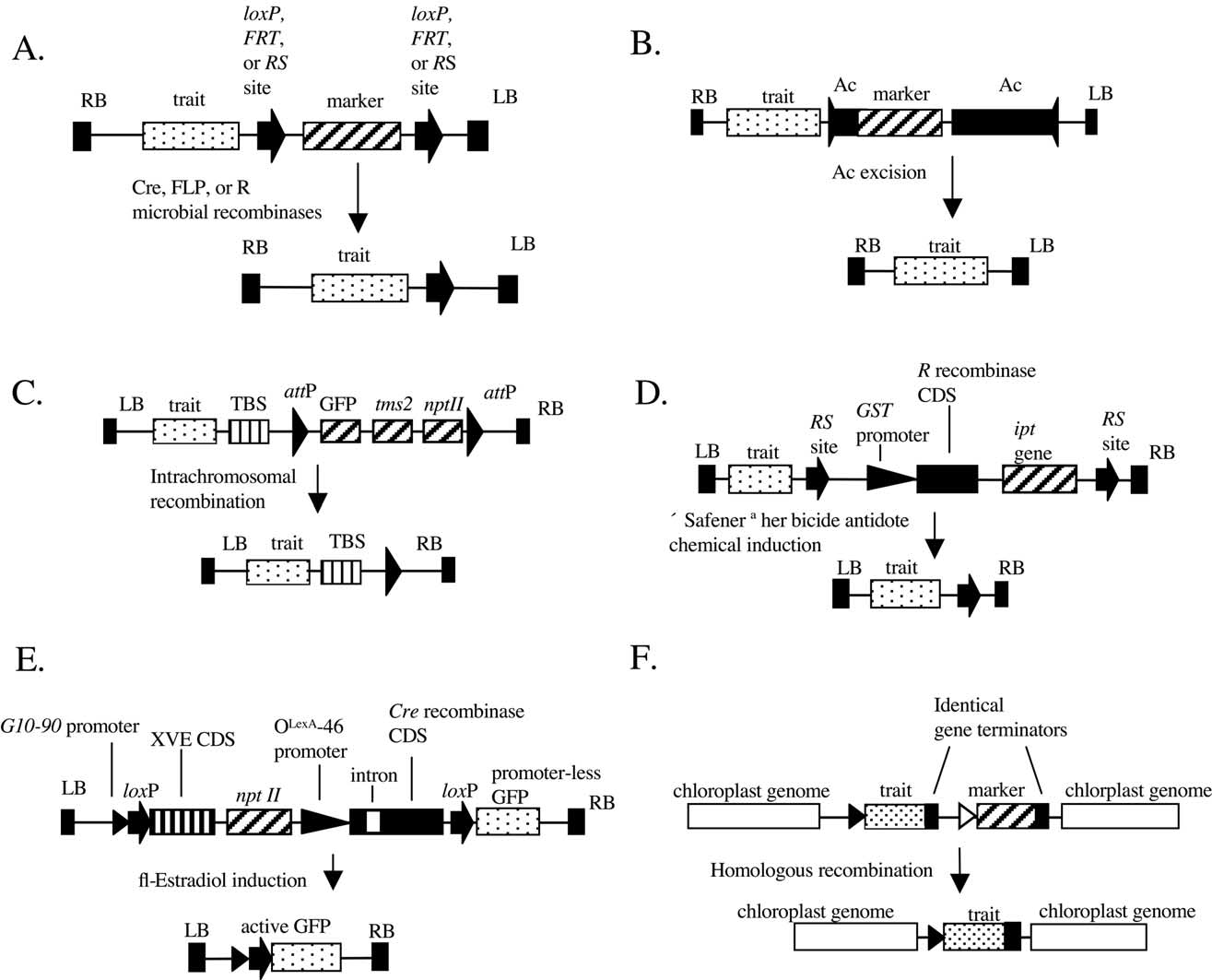 C. Scutt et al. / Biochimie 84 (2002) 1119–1126
Fig. 1. Transgene constructions used for the removal of marker genes from transgenic plants. (A) The simple use of microbial recombinases such as Cre, FLPand R (B) An Ac Transposon-based method for the removal of nuclear marker genes (C) The intrachromosomal recombination method for nuclearmarker gene removal (D) The GST-MAT vector system (E) The CRX vector system (F) Removal of chloroplast marker genes by homologousrecombination Ac = maize Activator transposable element, CDS = coding sequence, GFP = gene coding Green Fluorescent Protein. GST promoter =glutathione S-tranferase promoter, ipt = isopentyl transferase gene, LB and RB = left and right Agrobacterium tumefaciens T-DNA border sequences, nptII =neomycin phosphotransferase II gene, TBS = transformation booster sequence, tms2 = indolacetamide amidohydrolase gene, XVE = estrogen-activated hybridtranscriptional regulator gene
removed from such plants by genetic segregation. Co-
Zubko et al. tested the efficiency of a pair of 352 bp attP
transformation methods suffer from the obvious inefficiency
regions from bacteriophage k as substrates for ICR in plants.
C. Scutt et al. / Biochimie 84 (2002) 1119–1126
Fig. 1. Transgene constructions used for the removal of marker genes from transgenic plants. (A) The simple use of microbial recombinases such as Cre, FLPand R (B) An Ac Transposon-based method for the removal of nuclear marker genes (C) The intrachromosomal recombination method for nuclearmarker gene removal (D) The GST-MAT vector system (E) The CRX vector system (F) Removal of chloroplast marker genes by homologousrecombination Ac = maize Activator transposable element, CDS = coding sequence, GFP = gene coding Green Fluorescent Protein. GST promoter =glutathione S-tranferase promoter, ipt = isopentyl transferase gene, LB and RB = left and right Agrobacterium tumefaciens T-DNA border sequences, nptII =neomycin phosphotransferase II gene, TBS = transformation booster sequence, tms2 = indolacetamide amidohydrolase gene, XVE = estrogen-activated hybridtranscriptional regulator gene
removed from such plants by genetic segregation. Co-
Zubko et al. tested the efficiency of a pair of 352 bp attP
transformation methods suffer from the obvious inefficiency
regions from bacteriophage k as substrates for ICR in plants.