Do you want to buy antibiotics online without prescription? https://buyantibiotics24h.net/ - This is pharmacy online for you!
Ebd06.ebd.csic.es
Variation in herbivory by Yponomeuta mahalebella on
its only host plant Prunus mahaleb along an
C O N C H I T A A L O N S O EstacioÂn BioloÂgica DonÄana, C.S.I.C., Sevilla, Spain
Abstract. 1. The effect of natural variation in abiotic conditions on the herbivory
interaction between Prunus mahaleb (Rosaceae) and its monophagous folivore,
larvae of Yponomeuta mahalebella (Lepidoptera, Yponomeutidae), was analysed for
2 consecutive years along an elevational gradient in Sierra de Cazorla, south-east
2. There was a negative correlation between site elevation and mean population
herbivory level measured at the end of the growing period of Y. mahalebella.
Mortality during larval development was higher at higher elevation sites, and mean
adult body mass was higher in lower elevation populations.
3. Variation in temperatures recorded during the larval growth period at different
altitudes was the only study factor related to abundance of Y. mahalebella larvae;
neither differences in parasitisation rates nor plant features covaried signi®cantly
4. These results support the existence of geographical variation in plant±animal
interactions in relation to environmental heterogeneity. Key words. Elevational gradient, herbivory, Lepidoptera, natural enemies, plant±
insect interaction, temperature, Yponomeutidae.
dinal range distribution of P. mahaleb at Sierra de Cazorla,
JaeÂn province, south-east Spain (37°59¢N, 2°54¢W). Prunus
Thompson's (1994) review of plant±animal interactions
mahaleb is a small deciduous tree (2±8 m tall) of central and
pointed out that these interspeci®c relationships can be
southern Europe that, within the Sierra de Cazorla, normally
modi®ed by environmental conditions (e.g. Janzen, 1985;
grows in small groups of less than 10 individuals, with a few
Dudt & Shure, 1994; Herrera, 1995a,b and references therein;
scattered larger populations. Leaf ¯ush begins in mid-April
Louda & Rodman, 1996). Studies of plant±animal interactions
and development continues to the end of May. There is an
along natural abiotic gradients, e.g. latitudinal and altitudinal
approximately 10-day delay in ¯ushing between populations
(McCoy, 1990; Stevens, 1992; Begon et al., 1996; Gaston
at different elevations within the range considered in this
et al., 1998), can help to evaluate the role of environmental
study (C. Alonso, unpublished). The leaves of P. mahaleb
factors in such relationships. Such studies are, however, scarce
contain coumarins and hydroxicoumarins (Fung & Herreb-
(e.g. Koptur, 1985; Hill & Hodkinson, 1992), and conducted
out, 1987) that may play a feeding-deterrent role for many
mainly in extreme climates, such as the Arctic, where abiotic
factors are limiting for most species (Strathdee & Bale, 1998).
In late summer, females of Y. mahalebella deposit egg
More general conclusions may be obtained by studying
masses around the stems of P. mahaleb (Kooi, 1990). Eggs
altitudinal gradients in less extreme conditions.
hatch before hibernation and ®rst-instar larvae remain inside
This study focuses on the highly speci®c relationship
the batch shelter until the following spring. Caterpillars ®rst
between the Saint Lucie's Cherry (Prunus mahaleb L.,
become active in early April at lower sites and development
Rosaceae) and the small ermine moth (Yponomeuta mahale-
continues until late June. From the second instar on, larvae,
bella Latr., Lepidoptera: Yponomeutidae), within the altitu-
which are monophagous on P. mahaleb leaves, spin a
conspicuous silk tent and feed gregariously on leaves therein.
Correspondence: Conchita Alonso, Section of Ecology, Department
When food within the tent becomes limiting, caterpillars
of Biology, University of Turku, FIN-20014, Turku, Finland. E-mail:
extend the tent along the same branch. The cylindrical tents
can be measured easily in the ®eld.
The work reported here aimed to investigate which factors
the largest P. mahaleb populations located mainly between
affect the study plant±herbivore system and particularly
whether abiotic factors may be relevant for herbivory
Observations were carried out during the period in which Y.
interactions in a temperate area. Patterns of covariation
mahalebella larvae develop, from April to June, in 1994 and
between air temperature, herbivory level, and herbivore
1995. All P. mahaleb trees at each study population were
performance were studied over 2 consecutive years in six P.
marked initially in 1994 (see Table 1), except at the Torcal del
mahaleb populations located at different elevations in the study
Cerecino and Nava las Correhuelas sites, where only a
area. Variations in the impact of natural enemies and plant
subsample was marked owing to the large number of
features were also considered as alternative explanations to
individuals present. At the same time, a minimum±maximum
differences in herbivory along the study abiotic gradient. The
thermometer was attached to the shaded trunk of one tree at
speci®c questions addressed were: (1) How are Y. mahalebella
each population, and minimum and maximum weekly
larvae distributed among P. mahaleb populations? (2) Is their
temperatures were recorded over the study period (Table 1).
abundance related to site elevation? (3) Can natural variation
Mean weekly temperature was computed as (maximum +
in temperature, plant features, or natural-enemy populations
Marked trees were examined weekly and all Y. mahalebella
tents found were marked with numbered, coloured tape. In
1994, numbers of Y. mahalebella larvae within tents were
At the end of the season, total lengths of silk tents spun by Y.
Six P. mahaleb populations with altitudes ranging from 1300
mahalebella larvae were estimated for each tree. The length of
to 1700 m (Table 1), and distances between sites varying from
all individual tents in the tree, both with and without larvae
2 to 8 km, were selected in the southern part of Sierra de
inside, was summed. The incidence of Y. mahalebella at each
Cazorla Natural Park, a 214 000 ha protected area in south-east
site was calculated as the mean number of tents and mean tent
Spain (Fig. 1). The elevation of study populations was
determined directly by using an altimeter; however, because
Finally, to evaluate differences in herbivore ®tness among
the exact elevation for all P. mahaleb populations in the area
populations, all tents holding live larvae were collected
was not available, altitudinal distribution of P. mahaleb
between 21 and 24 June 1994, and 18 and 20 June 1995, by
(Fig. 2) was obtained by cartographic analysis, using a
clipping the branch portion they occupied. Branches were
1 : 50 000 scale map and detailed distribution information
placed in polythene bags and kept at room temperature until
(Fig. 1; C. M. Herrera and P. Jordano, unpublished). A map of
emergence of moths or parasitoids. Moths were killed, sexed,
the area was reticulated and the average elevation for every
dried at room temperature, and weighed. Parasitoid incidence
1-km2 area where P. mahaleb was present (n = 127, Fig. 1) was
was evaluated in collected samples as the proportion of
calculated as the mean of their minimum and maximum
parasitised individuals per tent. Although there are some
elevations. An altitudinal pro®le of the southern half of the
methodological problems with estimation of parasitoid in-
park where P. mahaleb is mainly located (Fig. 1) was drawn
cidence from ®eld collections (Van Driesche et al., 1991), it is
using the same method (i.e. calculating the mean of minimum
unlikely that parasitised larvae changed behaviour, because
and maximum elevation per 1 km2), and choosing a similar
this has not been reported for the same parasitoid species in
number of squares randomly (n = 124). This random sample
other Yponomeuta sp. (Dijkerman et al., 1986; Kuhlman,
represented 10% of the study area. This procedure indicated
1995). Parasitism rate data should be therefore treated with
that P. mahaleb distribution in the area ranges from 900 to
caution, but they were suitable for comparisons among
1900 m, with 50% of locations between 1400 and 1650 m, and
Table 1. Abiotic conditions of studied populations: elevation, and maximum, minimum, and mean weekly temperatures recorded during the 10-
week study period. Note that the differences among populations in elevation are not of the same magnitude, with two low, two intermediate, and
two high elevation sites. n = number of marked trees. Mean temperature was calculated as (maximum + minimum)/2.
# 1999 Blackwell Science Ltd, Ecological Entomology, 24, 371±379
Herbivory along an elevational gradient 373
for 100 tents collected in late June 1994. The relationship
between number of larvae and length of the tent was also
Length of Y. mahalebella tents was used as an estimate
studied experimentally. On 2 June 1995, ®ve tents were
of herbivore consumption and abundance. To test the
collected from an unmarked tree. Caterpillars from each
accuracy of this measurement, length of the tent, the
tent were taken out of the tent and divided in three groups,
number of caterpillars (herbivore load), and the number of
each with double the number of larvae of the previous
leaves consumed (herbivory level) within it, were recorded
group (e.g. 8±16±32). Fifteen branches from another
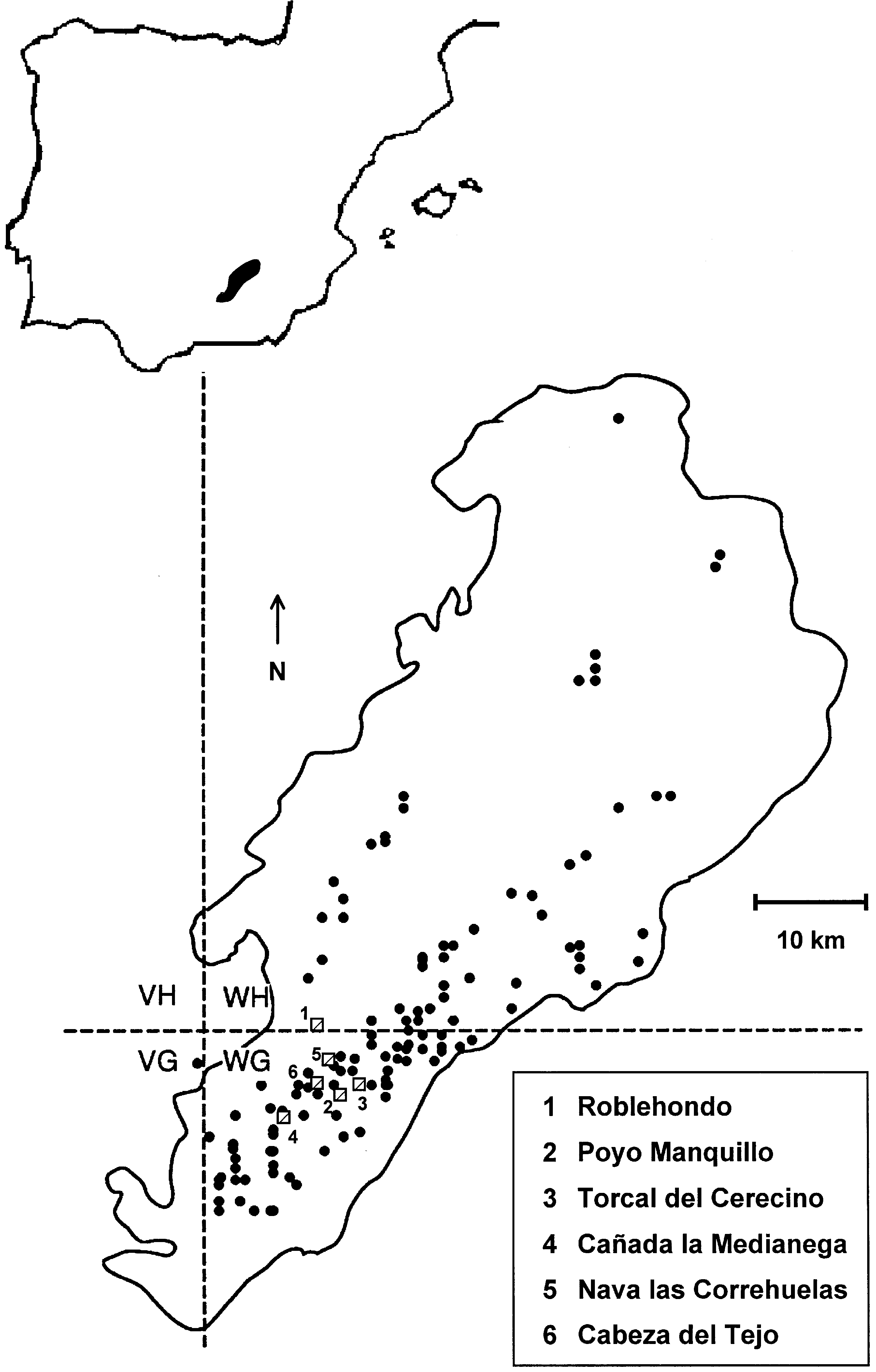
Fig. 1. Map of the study area showing the location of Sierra de Cazorla Natural Park, the distribution of P. mahaleb populations within it (dots),
and the study sites (dashed squares).
# 1999 Blackwell Science Ltd, Ecological Entomology, 24, 371±379
All analyses, except those stated above, were conducted
using the SAS package (SAS Institute, 1996). Means are
Length of the tent in centimetres (X) was related directly to
the number of eaten leaves within the tent (Y = 1.8 X); the
relationship was highly signi®cant (F = 242.7, d.f. = 1,98,
P < 0.001) and the regression explained a high percentage of
variance (R2 = 0.92). Tent length (X) was also related
signi®cantly to the number of caterpillars present at the end of
the growing season (Y = 1.25 + 0.21 X; F = 40.8, d.f. = 1,97,
Fig. 2. Altitudinal distribution of P. mahaleb in the Sierra de
P < 0.001), however this model explained a lower percentage of
Cazorla Natural Park. Black bars represent the percentage of P.
variance (R2 = 0.26). For this reason, the relationship was also
mahaleb populations (n = 127) at different altitudes, with grey areas
studied using a manipulative experiment where the number of
showing populations with > 10 individuals. Dashed ®gure represents
caterpillars present was the only factor related signi®cantly to
the percentage of land area at different altitudes, estimated from a
the length of the tent after 5 days (F = 41.6, d.f. = 1,5, P = 0.001).
random sample of 1-km2 squares (n = 124) within the area (see
Number of larvae explained a high percentage of variance in
tent length (R2 = 0.83) in a model including also origin of the
larvae and origin 3 number interaction.
At tree level, the total tent length was correlated signi®-
cantly with the number of tents [rs (1994) = 0.88, n = 122,
unmarked tree, each » 25 cm long, were cut and put into
P < 0.001; rs (1995) = 0.98, n = 122, P < 0.001].
clip vials. Each group of caterpillars was put onto a branch
portion and covered with transparent plastic bags to prevent
escape. Branches were kept at room temperature, and the
Among-populations variation in air temperature
length of tents was measured after 5 days.
Size of the trees did not affect the estimates at tree level
Maximum, minimum, and mean weekly temperatures
because larvae were never constrained by scarcity of free space
recorded between mid-April and late-June did not differ
signi®cantly from 1994 to 1995 (d.f. = 1,109, P = NS, for three
separate ANOVAs; Table 1). Both maximum and minimum
temperatures differed signi®cantly among sites [F (maxi-
mum) = 3.1, d.f. = 5,105, P < 0.05; F (minimum) = 3.2,
d.f. = 5,105, P < 0.05]. Maximum temperatures at the CanÄada
Differences in herbivory level among populations were
la Medianega site were signi®cantly lower than at the other
evaluated using the nonparametric Kruskal±Wallis test,
study sites, and minimum temperatures were signi®cantly
because neither the number of tents nor the total tent length
lower at Nava las Correhuelas than at the other sites. Mean
temperatures were higher at the two low elevation sites than at
The relationships between tent length and both herbivory
the highest elevations (Table 1), but differences among sites in
level and herbivore load were analysed by robust regression
mean temperature were not statistically signi®cant (F = 2.05,
using the least median squared method within the PROGRESS
program (Rousseeuw & Leroy, 1987).
When there was an ordered prediction about differences
among populations, isotonic regression was used (Gaines &
Differences in herbivory among populations
Finally, tents were used as sample units to evaluate
Study populations differed signi®cantly in herbivory level [c2
differences in herbivore performance among populations.
(1994) = 40.85, d.f. = 5, P < 0.001; c2 (1995) = 46.11, d.f. = 5,
Caterpillars growing within the same tent were not indepen-
P < 0.001, Kruskal±Wallis tests]. Although overall herbivory
dent, probably belonging to the same brood, hatching at the
was higher in 1994, differences among populations were
same place, and growing in the same micro-environment, and
consistent in both study seasons (rs = 0.94, n = 6, P < 0.01;
thus were not statistically independent. The percentage of
collected larvae reaching the adult stage was used to estimate
In 1994, herbivory at the population level was correlated
survival, and adult body mass was used as an estimate of
negatively with site altitude (rs = ± 0.83, n = 6, P < 0.05) and
positively with mean temperature recorded during the study
# 1999 Blackwell Science Ltd, Ecological Entomology, 24, 371±379
Herbivory along an elevational gradient 375
Fig. 3. Mean population herbivory level observed in the six P.
mahaleb study populations in 1994 and 1995. Symbols represent
mean values, bars = ±SEM, and the dotted line represents equality
between years. Populations are ordered by altitude from lowest
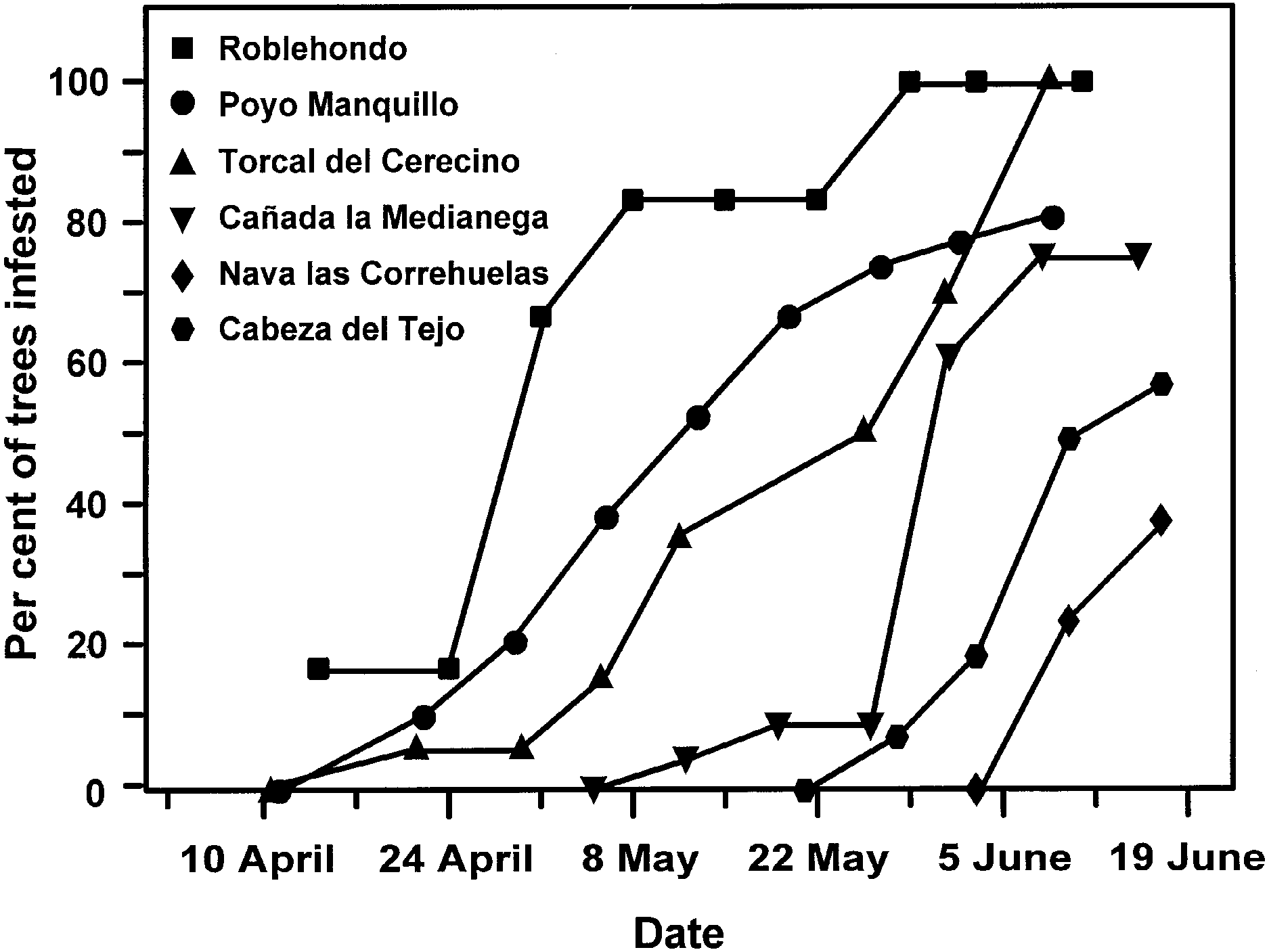
Fig. 5. Phenology of Y. mahalebella tent appearance at different
(Roblehondo) to highest (Cabeza del Tejo) elevation.
locations, showing the percentage of trees at every location where
there was at least one Y. mahalebella tent during the 10-week study
period. Note that all trees presented Y. mahalebella tents at the end
of the season in only two populations. Populations are ordered by
altitude from lowest (Roblehondo) to highest (Cabeza del Tejo)
lower altitude populations, but they reached the last instar
simultaneously at all sites, indicating longer growth periods at
Adult body mass. Adult body mass was used as an estimate
of potential fecundity. As noted above, larvae within the same
tent could not be treated as independent samples, so analyses
were conducted after computing means for individual tents and
using numbers of individuals per tent as a weighting factor
(SAS Institute, 1996). Females were signi®cantly heavier
(7.72 6 1.98 mg, n = 289)(F = 233.56, d.f. = 1,532, P < 0.001).
Mean adult body mass per tent was therefore calculated
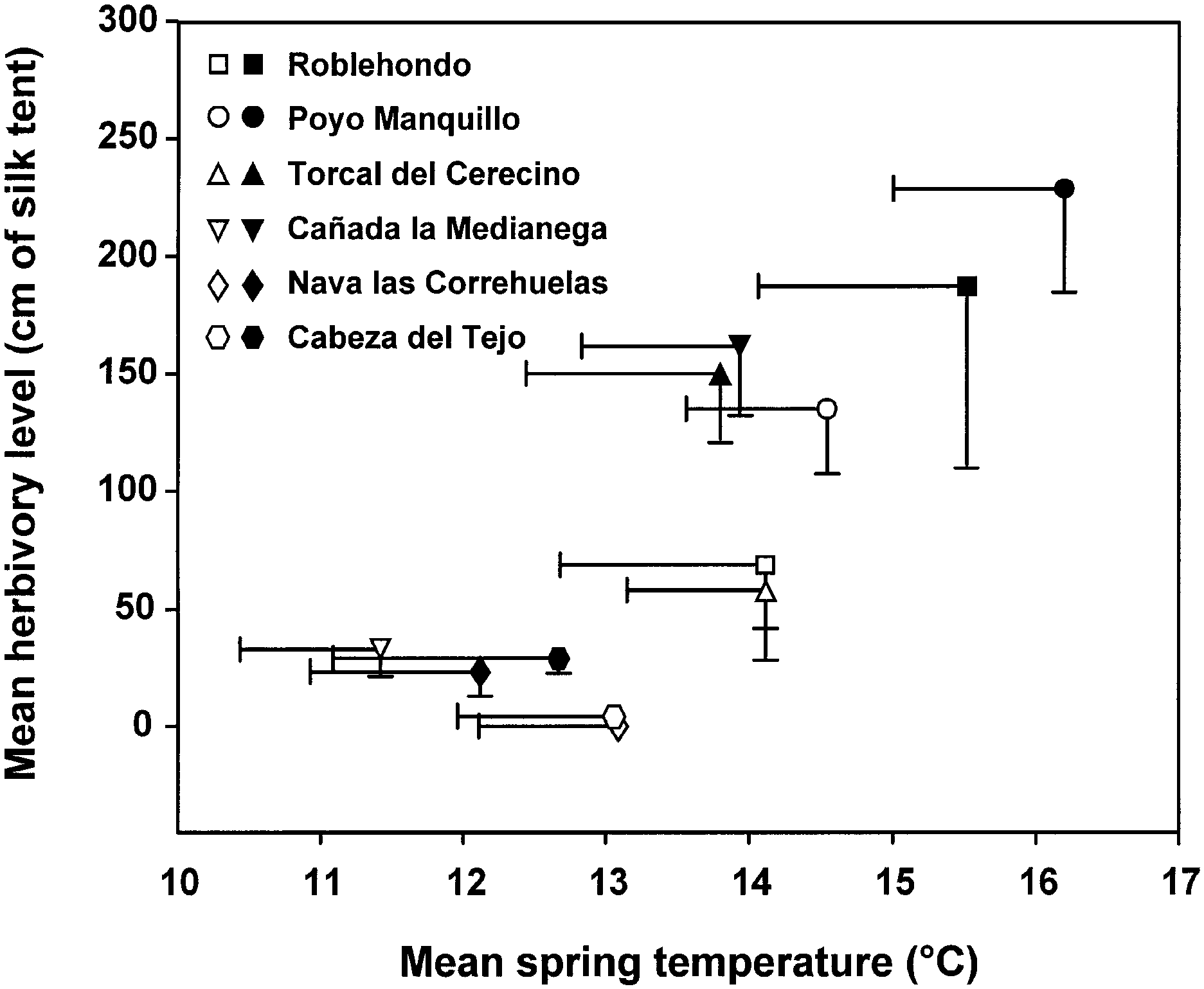
Fig. 4. Relationship between mean population herbivory level and
Effects of sex, year, population, and year 3 population
mean temperature recorded during the 10-week study period (April±
factors on adult body mass were evaluated. All factors
June) of Y. mahalebella larval development. Symbols represent mean
except sex were considered as random effects in a general
values, and bars = ±SEM. Different symbols identify study
linear model (Proc GLM; SAS Institute, 1996). The
populations, with black symbols representing 1994 data and white
symbols representing 1995 data. Populations are ordered by altitude
d.f. = 11,145, P < 0.001) and explained 54% of variance in
from lowest (Roblehondo) to highest (Cabeza del Tejo) elevation.
adult body mass. Neither year (F = 0.11, d.f. = 1,8.58,
P = NS) nor population (F = 0.76, d.f. = 5,4.12, P = NS)
showed a signi®cant effect on adult body mass (Table 2),
season (rs = 1, n = 6, P < 0.01; Fig. 4). This pattern was
but there was a signi®cant year 3 population interaction
supported by the 1995 results (isotonic regression:
(F = 4.03, d.f. = 4,145, P < 0.01) in addition to the sex effect.
E2 = 0.273, n = 6, P < 0.001), where mean population herbivory
Separating the data by year, population had a signi®cant
increased steadily with mean population temperature.
effect in both data sets [F (1994) = 3.08, d.f. = 5,72,
P < 0.05; F (1995) = 6.47, d.f. = 4,72, P < 0.001].
The relationships between adult body mass and both
Consequences for herbivores growing within different tree
population temperature and elevation were evaluated sepa-
rately for each study season owing to the signi®cant
year 3 population interaction in the general linear model (see
Phenology. Date of Y. mahalebella tent appearance was
above). In 1994, there was a positive relationship between
correlated with site altitude (Fig. 5). Tents appeared earlier in
adult body mass and mean temperature (Fig. 6), with a
# 1999 Blackwell Science Ltd, Ecological Entomology, 24, 371±379
Table 2. Adult body mass of Y. mahalebella individuals reared from ®nal-instar larvae collected from different populations. Mean population
dry body mass 6 SD (mg) is presented separately by sex and year. Population means were calculated based on means per tent and weighted for
number of individuals within each tent. Number of tents (nt) and number of individuals (ni) are shown. Missing values correspond to absence of
(F = 1.17, d.f. = 3,43, P = NS), however the ®nal numbers
differed signi®cantly among populations (F = 9.59, d.f. = 3,43,
P < 0.001; Table 3). The percentage of lost larvae during the
growing period was also signi®cantly different among
populations (F = 8.77, d.f. = 3,43, P < 0.001), with greater
losses recorded in higher altitude populations (Table 3),
however it was not possible to evaluate the effect of dispersal
Differences in parasitism rates among populations and
consistency between years were measured using data for the
three populations with adequate sample sizes in both study
years (Poyo Manquillo, Torcal del Cerecino, and CanÄada la
Medianega; Table 4). Differences among populations were
statistically signi®cant (generalised logits model: c2 = 11.06,
d.f. = 4, P < 0.05), with the parasitism rate lowest at Poyo
Manquillo site. The population 3 year interaction was also
signi®cant (c2 = 33.86, d.f. = 4, P < 0.001), indicating that there
were differences among populations in larval performance
Fig. 6. Relationship between mean adult Y. mahalebella body mass
and mean temperature recorded at different populations during the
larval development period. Symbols represent mean values, and
bars = ±SEM. Populations are ordered by altitude from lowest (Poyo
Manquillo) to highest (Cabeza del Tejo) elevation.
The incidence of herbivores on plants is usually estimated in
the literature as either plant defoliation or herbivore abun-
signi®cant steady increase of adult body mass at lower
dance, but only rarely have both kinds of measurements been
altitudes (isotonic regression E2 = 0.12, n = 5, P < 0.001 for
considered simultaneously (see Alonso & Herrera, 1996). The
females; E2 = 0.08, n = 5, P < 0.001 for males). A similar result
tight relationship found here between tent length and the
was found for 1995 data where only four populations could be
number of consumed leaves within it, along with the
used due to reduced sample sizes (E2 = 0.08, n = 4, P < 0.05 for
experimental evidence that tent length was highly dependent
females; E2 = 0.19, n = 4, P < 0.001 for males).
of the number of larvae spinning the tent, served to
demonstrate that the length of Y. mahalebella tents may be
Herbivore mortality. Two different aspects were considered:
used as a reliable indirect estimate of both defoliation and
larval disappearance during the growing season and parasitism.
The mean number of caterpillars per tent was signi®cantly
Estimated abundance of Y. mahalebella larvae was corre-
lower at the end of the growing season than at the beginning
lated inversely with site altitude over 2 successive years, a
(t = 7.9, n = 47, P < 0.001; Table 3). Initial mean number in
pattern supported by casual observations over many years
different populations varied between 19 and 24 caterpillars per
(C. M. Herrera, pers. comm.), indicating that P. mahaleb
tent, and did not differ signi®cantly among populations
populations above 1600 m never receive massive defoliations,
# 1999 Blackwell Science Ltd, Ecological Entomology, 24, 371±379
Herbivory along an elevational gradient 377
Table 3. Caterpillar disappearance during the 1994 growing season. ni is the initial estimated number of caterpillars within a tent, nf is the
number of caterpillars collected at the end of the growing season. Per cent loss was calculated from the difference between ni and nf in collected
tents. Different letters in the same column indicate statistically-signi®cant differences (Student±Newman±Keuls test, P < 0.05).
Table 4. Growth result of rearing ®nal-instar larvae collected from different populations. nt is the number of collected tents and nc the number
of collected caterpillars. Mean percentages of adults (A), parasitised (P), and dead (R) caterpillars were calculated based on individual tent data.
but that these are more frequent below 1400 m. Yponomeuta
warmer sites also have longer growing periods. Finally, adult
mahalebella was more abundant at lower elevations, where P.
moths at lower elevations were heavier (Fig. 6), in accord with
mahaleb populations were less well represented (Fig. 2), and
the model for spatial heterogeneity proposed by Sibly and
was uncommon at the middle and high elevations, where the
Atkinson (1994) in which `the optimal plastic strategy is larger
largest P. mahaleb populations were present (Fig. 2). This
adult body size¼in warmer habitats'. As body size is usually
suggests that there was some factor(s) affecting the species
linked to fecundity in Yponomeuta (Kooi et al., 1989; Leather
distributions differentially. A species such as P. mahaleb, at
& Mackenzie, 1994), individuals at lower elevations probably
the southern limit of its distribution, may be less stressed and
have higher fecundity, which contributes to their higher
therefore more abundant at higher elevations, where abiotic
conditions are more similar to those recorded at more northerly
Losses of Y. mahalebella larvae during development were
also fewer at the lower sites (Table 3). Climate unpredict-
Factors determining the elevational pattern of Y. mahale-
ability, plant phenology, and natural enemies could not
bella abundance were less evident. Many factors can affect
apparently predict the observed pattern of mortality. Myers
insect herbivore performance along an elevational gradient,
(1981) suggested that larvae commencing growth late in spring
including plant phenology (Hodkinson, 1997), plant secondary
will tend to have lower mortality because climate variability is
compounds (Louda & Rodman, 1983; Koptur, 1985), leaf traits
less than in early spring, however Y. mahalebella larval
(Kudo, 1996), and abundance of natural enemies (Koptur,
disappearance was highest in the upper populations, which
1985). In addition, variation in abiotic conditions related to
began to grow later (Fig. 5). More detailed studies are needed
elevation can similarly affect larval development and abun-
to evaluate whether small differences in synchrony between
dance. Finally, both biotic and abiotic factors may also vary
leaf ¯ush and the start of larval activity (Aide & LondonÄo,
among sites at similar elevations (Sork et al., 1993). The highly
1989) explain differences in disappearance rates, although on
signi®cant correlation found between herbivore abundance and
the coarse scale studied, phenology does not appear to explain
site temperature (Fig. 4) suggests that temperature in¯uences
differences in Y. mahalebella abundance. Tent appearance was
herbivore abundance at different altitudes, probably through
delayed by approximately 2 weeks at higher elevation sites
differences in degree-day accumulation (Whittaker & Tribe,
(Fig. 5), but because the host trees also ¯ushed leaves later,
1996; Hodkinson, 1997; RuohomaÈki et al., 1997 and references
interspeci®c synchronisation should not be affected unduly,
therein). Furthermore, differences in temperature among sites
especially because external folivores such as Y. mahalebella
may be ampli®ed by tent structure, because the temperature
larvae are unlikely to be constrained by a lack of suitable food
within tents is related to sun exposure (Alonso, 1997a), and the
# 1999 Blackwell Science Ltd, Ecological Entomology, 24, 371±379
Both parasitisation and predation rates also fail to explain
Alonso, C. (1997b) Variaciones en las relaciones planta-insectos
larval disappearance during development. Toxic compounds
®toÂfagos: efectos de factores bioÂticos y abioÂticos. PhD thesis,
contained in Y. mahalebella larvae (Fung & Herrebout,
1987; Fung, 1988), combined with gregariousness (SilleÂn-
Alonso, C. & Herrera, C.M. (1996) Variation in herbivory within and
Tullberg & Leimar, 1988; Gamberale & Tullberg, 1996)
among plants of Daphne laureola (Thymelaeaceae): correlation
and protection within tents all serve as predator deterrents.
with plant size and architecture. Journal of Ecology, 84, 495±502.
Begon, M., Harper, J.L. & Townsend, C.R. (1996) Ecology:
In addition, it might be expected that mortality would be
Individuals Populations and Communities, 3rd edn. Blackwell,
higher where larval growing periods were longer, giving
greater opportunity for predator attack (Evans, 1982; but
Benrey, B. & Denno, R.F. (1997) The slow-growth-high-mortality
see Benrey & Denno, 1997), but the observed pattern was
hypothesis: a test using the cabbage butter¯y. Ecology, 78, 987±
just the opposite. Finally, parasitoids were not expected to
affect mortality during the early developmental stages,
Dijkerman, H.J., Groot, J.M.B. & Herrebout, W.M. (1986) The
because the main parasitoid species, such as Ageniaspis
parasitoids of the genus Yponomeuta Latreille (Lepidoptera,
fuscicollis Dalman and Diadegma sp., emerge from ®nal-
Yponomeutidae) in the Netherlands. Proccedings of Kononklijke
instar larvae (see Kuhlman, 1995 and references therein).
Netherlandse Akademie Van Wetenschappen C, 84, 379±398.
Moreover, parasitism rates recorded from ®nal-instar larvae
Dudt, J.F. & Shure, D.J. (1994) The in¯uence of light and nutrients on
were higher in the Poyo Manquillo population where the
foliar phenolics and insect herbivory. Ecology, 75, 86±98.
highest herbivore abundances were recorded. Otherwise,
Evans, E.W. (1982) In¯uence of weather on predator/prey relations:
stinkbugs and tent caterpillars. New York Entomological Society, 90,
neither differences in plant size nor the mineral or physical
properties of P. mahaleb leaves were correlated signi®-
Fung, S.Y. (1988) Butenolides in parasitoids of small ermine moths,
cantly with recorded herbivore abundance in the study
Yponomeuta spp. (Lepidoptera: Yponomeutidae). Proceedings of
Koninklijke Netherlandse Akademie Van Wetenschappen C, 91,
The observed elevational pattern of Y. mahalebella abun-
dance therefore appears to be related mainly to variation in
Fung, S.Y. & Herrebout, W.M. (1987) Coumarins in Prunus mahaleb
temperature affecting fecundity. The relationship between
and its herbivore the small ermine moth Yponomeuta mahalebellus.
temperature and larval mortality remains unknown, but biotic
Journal of Chemical Ecology, 13, 2041±2047.
factors did not explain the lower mortality found at the lower
Gaines, S.D. & Rice, W.R. (1990) Analysis of biological data when
elevation sites. A detailed study of thermal requirements of Y.
there are ordered expectations. The American Naturalist, 135, 310±
mahalebella larvae will contribute to elucidating whether
temperature can also be related to larval mortality. Never-
Gamberale, G. & Tullberg, B.S. (1996) Evidence for a more effective
signal in aggregated aposematic prey. Animal Behaviour, 52, 597±
theless, these results suggest that the magnitude of herbivore
pressure can be modi®ed by environmental conditions within a
Gaston, K.J., Blackburn, T.M. & Spicer, J.I. (1998) Rapoport's rule:
geographical range (Thompson, 1994).
time for an epitaph? Trends in Ecology and Evolution, 13, 70±74.
Herrera, C.M. (1995a) Floral biology, microclimate and pollination by
ectothermic bees in an early-blooming herb. Ecology, 76, 218±228.
Herrera, C.M. (1995b) Microclimate and individual variation in
pollinators: ¯owering plants are more than their ¯owers. Ecology,
I am particularly indebted to Carlos M. Herrera for teaching,
encouragement and valuable advice. He, Jose M. GoÂmez,
Hill, J.K. & Hodkinson, I.D. (1992) Effects of temperature on
phenological synchrony and altitudinal distribution of jumping
Erkki Haukioja, Ian D. Hodkinson, Simon R. Leather, Lauri
plant lice (Hemiptera: Psylloidea) on dwarf willow (Salix
Oksanen and two anonymous reviewers made useful com-
lapponum) in Norway. Ecological Entomology, 20, 237±244.
ments on earlier versions of this manuscript. Manolo CarrioÂn,
Hodkinson, I.D. (1997) Progressive restriction of host plant exploita-
Alicia Prieto, and RocõÂo Requerey assisted with sampling and
tion along a climatic gradient: the willow psyllid Cacopsylla
processing data. The Agencia de Medio Ambiente supported
groenlandica in Greenland. Ecological Entomology, 22, 47±54.
this work in Cazorla, and Spanish DGICYT and Ministerio de
Hodkinson, I.D. & Bird, J. (1998) Host-speci®c insect herbivores as
EducacioÂn y Ciencia funded it through grant PB91±0114, and a
sensors of climate change in Arctic and Alpine environments. Arctic
predoctoral fellowship. While writing the paper I was
supported by a European Commission-TMR fellowship.
Janzen, D.H. (1985) A host plant is more than its chemistry. Illinois
Natural History Survey Bulletin, 33, 141±174.
Kooi, R.E. (1990) Host-plant selection and larval food-acceptance by
small ermine moths. PhD thesis, University of Leiden. Studies in
Yponomeuta, 9. ICG Printing, Dordrecht, The Netherlands.
Kooi, R.E., Herrebout, W.M. & van de Water, T.P.M. (1989)
Aide, T.M. & LondonÄo, E.C. (1989) The effects of rapid leaf expansion
Observations on pupal and imaginal weights of small ermine moths
on the growth and survivorship of a lepidopteran herbivore. Oikos,
(Lepidoptera: Yponomeuta). Proceedings of the Koninklijke Neder-
landse Akademie Van Wetenschappen C, 92, 81±88.
Alonso, C. (1997a) Choosing a place to grow. Importance of within-
Koptur, S. (1985) Alternative defenses against herbivores in Inga
plant abiotic microenvironment for Yponomeuta mahalebella.
(Fabaceae: Mimosidae) over an elevational gradient. Ecology, 66,
Entomologia experimentalis et applicata, 83, 171±180.
# 1999 Blackwell Science Ltd, Ecological Entomology, 24, 371±379
Herbivory along an elevational gradient 379
Kudo, G. (1996) Intraspeci®c variation of leaf traits in several
SAS Institute (1996) SAS/STAT Software: Changes and Enhancements
deciduous species in relation to length of growing season.
through Release 6.11. SAS Institute, Cary, North Carolina.
Sibly, R.M. & Atkinson, D. (1994) How rearing temperature affects
Kuhlman, U. (1995) Impact of insect predators and parasitoids on
optimal adult size in ectotherms. Functional Ecology, 8, 486±493.
populations of the apple ermine moth, Yponomeuta malinellus,
SilleÂn-Tullberg, B. & Leimar, O. (1988) The evolution of gregarious-
(Lepidoptera: Yponomeutidae). PhD thesis, International Institute of
ness in distasteful insects as a defense against predators. The
Biological Control, DeleÂmont, Switzerland.
Leather, S.R. & Mackenzie, G.A. (1994) Factors affecting the
Sork, V., Stowe, K.A. & Hochwender, C. (1993) Evidence for local
population development of the bird cherry ermine moth, Ypono-
adaptation in closely adjacent populations of northern red oak
meuta evonymella (L.). The Entomologist, 113, 86±105.
(Quercus rubra L.) expressed as resistance to leaf herbivores. The
Louda, S.M. & Rodman, J.E. (1983) Ecological patterns in the
glucosinolate content of a native mustard, Cardamine cordifolia, in
Stevens, G.C. (1992) The elevational gradient in altitude range: an
the Rocky Mountains. Journal of Chemical Ecology, 9, 397±422.
extension of the Rapoport's latitudinal rule of altitude. The
Louda, S.M. & Rodman, J.E. (1996) Insect herbivory as a major factor
in the shade distribution of a native crucifer (Cardamine cordifolia
Strathdee, A.T. & Bale, J.S. (1998) Life on the edge: insect ecology in
A. Gray, bittercress). Journal of Ecology, 84, 229±237.
arctic environments. Annual Review of Entomology, 43, 85±106.
McCoy, E.D. (1990) The distribution of insects along elevational
Thompson, J.N. (1994) The Co-evolutionary Process. University of
Myers, J.H. (1981) Interactions between western tent caterpillars and
Van Driesche, R.G., Bellows, T.S., Elkinton, J.S., Gould, J.R. & Ferro,
wild rose: a test of some general plant herbivore hypotheses.
D.N. (1991) The meaning of percentage parasitism revisited:
Journal of Animal Ecology, 50, 11±25.
solutions to the problem of accurately estimating total losses from
Rousseeuw, P.J. & Leroy, A.M. (1987) Robust Regression and Outlier
parasitism. Environmental Entomology, 20, 1±7.
Detection. Wiley-Interscience, New York.
Whittaker, J.B. & Tribe, N.P. (1996) An altitudinal transect as an
RuohomaÈki, K., Virtanen, T., Kaitaniemi, P. & Tammaru, T. (1997)
indicator of responses of spittlebug (Auchenorrhyncha: Cercopidae)
Old mountain birches at high altitudes are prone to outbreaks of
to climate change. European Journal of Entomology, 93, 319±324.
Epirrita autumnata (Lepidoptera: Geometridae). Environmental
# 1999 Blackwell Science Ltd, Ecological Entomology, 24, 371±379
Position Paper - Alcohol and other Drugs August 2013 The Inner South Community Health Service (ISCHS) is a major provider of primary health care services including alcohol and other drug (AOD) services, and has been for over 20 years. The service model is based on the social model of health. Principles of harm minimization are adopted in relation to all AOD work. 1. Purpose This
~~NEW SUMMER EDITION~~ Message from the President Barrett P. Brenton (St. John’s University) It is with great pleasure that I welcome NEAA Newsletter readers to the inaugural summer winner Erica Varlese from Drew University issue. The idea of an additional issue was and her exploration of labor support and devised under the leadership of Editor-in-resistance in American bir
 Herbivory along an elevational gradient 373
for 100 tents collected in late June 1994. The relationship
between number of larvae and length of the tent was also
Length of Y. mahalebella tents was used as an estimate
studied experimentally. On 2 June 1995, ®ve tents were
of herbivore consumption and abundance. To test the
collected from an unmarked tree. Caterpillars from each
accuracy of this measurement, length of the tent, the
tent were taken out of the tent and divided in three groups,
number of caterpillars (herbivore load), and the number of
each with double the number of larvae of the previous
leaves consumed (herbivory level) within it, were recorded
group (e.g. 8±16±32). Fifteen branches from another
Fig. 1. Map of the study area showing the location of Sierra de Cazorla Natural Park, the distribution of P. mahaleb populations within it (dots),
and the study sites (dashed squares).
Herbivory along an elevational gradient 373
for 100 tents collected in late June 1994. The relationship
between number of larvae and length of the tent was also
Length of Y. mahalebella tents was used as an estimate
studied experimentally. On 2 June 1995, ®ve tents were
of herbivore consumption and abundance. To test the
collected from an unmarked tree. Caterpillars from each
accuracy of this measurement, length of the tent, the
tent were taken out of the tent and divided in three groups,
number of caterpillars (herbivore load), and the number of
each with double the number of larvae of the previous
leaves consumed (herbivory level) within it, were recorded
group (e.g. 8±16±32). Fifteen branches from another
Fig. 1. Map of the study area showing the location of Sierra de Cazorla Natural Park, the distribution of P. mahaleb populations within it (dots),
and the study sites (dashed squares).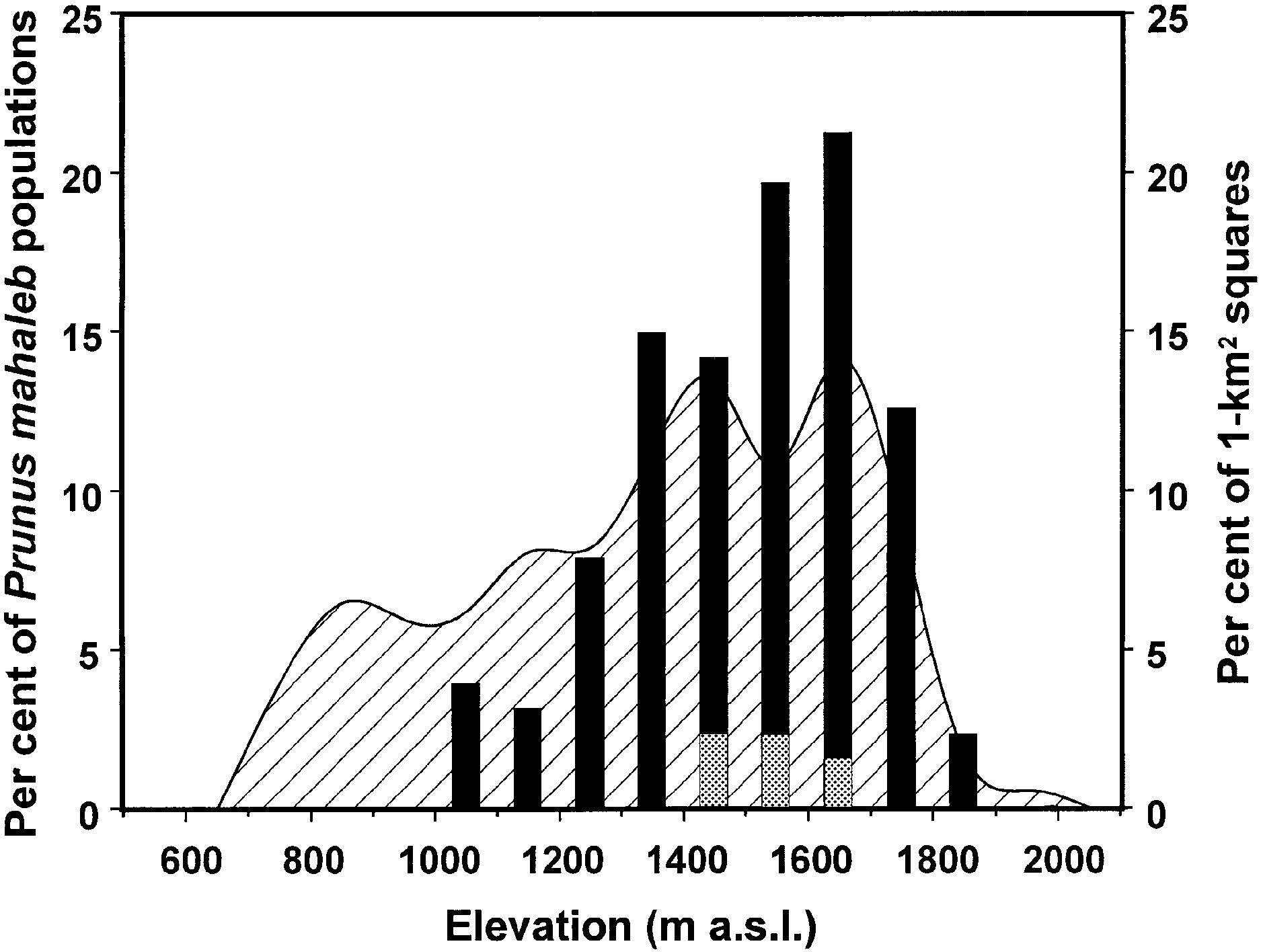 All analyses, except those stated above, were conducted
using the SAS package (SAS Institute, 1996). Means are
Length of the tent in centimetres (X) was related directly to
the number of eaten leaves within the tent (Y = 1.8 X); the
relationship was highly signi®cant (F = 242.7, d.f. = 1,98,
P < 0.001) and the regression explained a high percentage of
variance (R2 = 0.92). Tent length (X) was also related
signi®cantly to the number of caterpillars present at the end of
the growing season (Y = 1.25 + 0.21 X; F = 40.8, d.f. = 1,97,
Fig. 2. Altitudinal distribution of P. mahaleb in the Sierra de
P < 0.001), however this model explained a lower percentage of
Cazorla Natural Park. Black bars represent the percentage of P.
All analyses, except those stated above, were conducted
using the SAS package (SAS Institute, 1996). Means are
Length of the tent in centimetres (X) was related directly to
the number of eaten leaves within the tent (Y = 1.8 X); the
relationship was highly signi®cant (F = 242.7, d.f. = 1,98,
P < 0.001) and the regression explained a high percentage of
variance (R2 = 0.92). Tent length (X) was also related
signi®cantly to the number of caterpillars present at the end of
the growing season (Y = 1.25 + 0.21 X; F = 40.8, d.f. = 1,97,
Fig. 2. Altitudinal distribution of P. mahaleb in the Sierra de
P < 0.001), however this model explained a lower percentage of
Cazorla Natural Park. Black bars represent the percentage of P.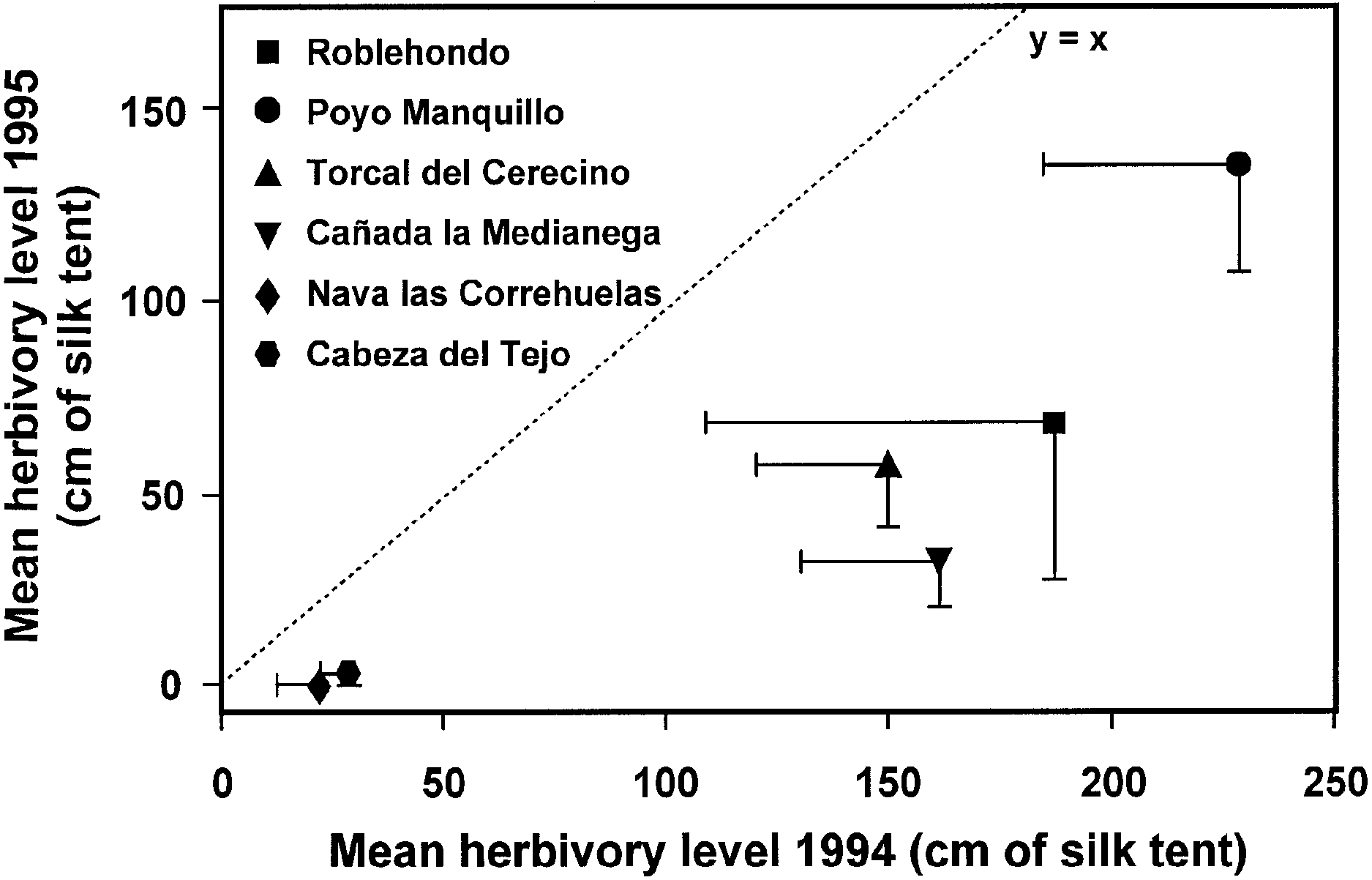
 Herbivory along an elevational gradient 375
Fig. 3. Mean population herbivory level observed in the six P.
Herbivory along an elevational gradient 375
Fig. 3. Mean population herbivory level observed in the six P.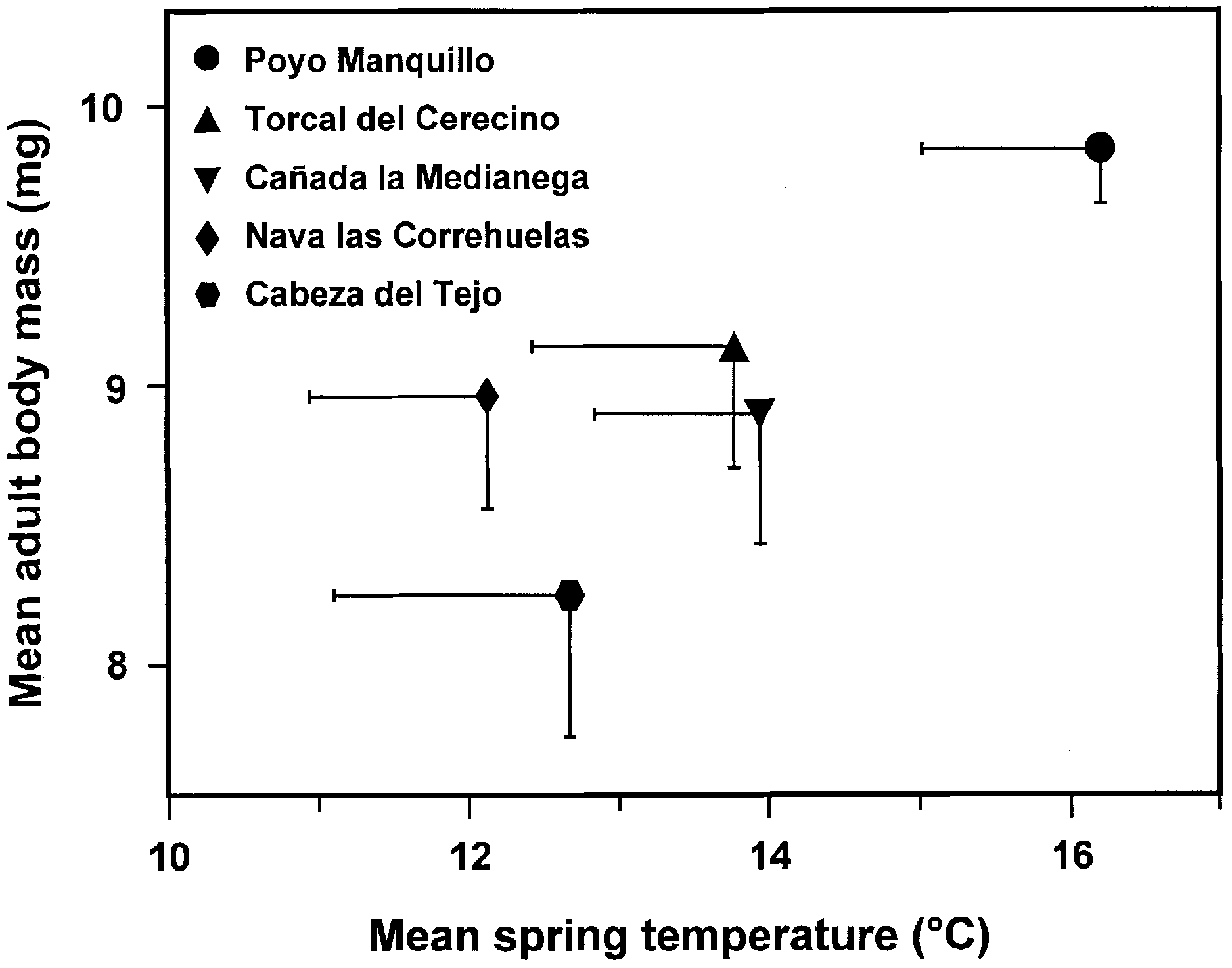 Table 2. Adult body mass of Y. mahalebella individuals reared from ®nal-instar larvae collected from different populations. Mean population
dry body mass 6 SD (mg) is presented separately by sex and year. Population means were calculated based on means per tent and weighted for
number of individuals within each tent. Number of tents (nt) and number of individuals (ni) are shown. Missing values correspond to absence of
(F = 1.17, d.f. = 3,43, P = NS), however the ®nal numbers
differed signi®cantly among populations (F = 9.59, d.f. = 3,43,
P < 0.001; Table 3). The percentage of lost larvae during the
growing period was also signi®cantly different among
populations (F = 8.77, d.f. = 3,43, P < 0.001), with greater
losses recorded in higher altitude populations (Table 3),
however it was not possible to evaluate the effect of dispersal
Differences in parasitism rates among populations and
consistency between years were measured using data for the
three populations with adequate sample sizes in both study
years (Poyo Manquillo, Torcal del Cerecino, and CanÄada la
Medianega; Table 4). Differences among populations were
statistically signi®cant (generalised logits model: c2 = 11.06,
d.f. = 4, P < 0.05), with the parasitism rate lowest at Poyo
Manquillo site. The population 3 year interaction was also
signi®cant (c2 = 33.86, d.f. = 4, P < 0.001), indicating that there
were differences among populations in larval performance
Fig. 6. Relationship between mean adult Y. mahalebella body mass
and mean temperature recorded at different populations during the
larval development period. Symbols represent mean values, and
bars = ±SEM. Populations are ordered by altitude from lowest (Poyo
Manquillo) to highest (Cabeza del Tejo) elevation.
Table 2. Adult body mass of Y. mahalebella individuals reared from ®nal-instar larvae collected from different populations. Mean population
dry body mass 6 SD (mg) is presented separately by sex and year. Population means were calculated based on means per tent and weighted for
number of individuals within each tent. Number of tents (nt) and number of individuals (ni) are shown. Missing values correspond to absence of
(F = 1.17, d.f. = 3,43, P = NS), however the ®nal numbers
differed signi®cantly among populations (F = 9.59, d.f. = 3,43,
P < 0.001; Table 3). The percentage of lost larvae during the
growing period was also signi®cantly different among
populations (F = 8.77, d.f. = 3,43, P < 0.001), with greater
losses recorded in higher altitude populations (Table 3),
however it was not possible to evaluate the effect of dispersal
Differences in parasitism rates among populations and
consistency between years were measured using data for the
three populations with adequate sample sizes in both study
years (Poyo Manquillo, Torcal del Cerecino, and CanÄada la
Medianega; Table 4). Differences among populations were
statistically signi®cant (generalised logits model: c2 = 11.06,
d.f. = 4, P < 0.05), with the parasitism rate lowest at Poyo
Manquillo site. The population 3 year interaction was also
signi®cant (c2 = 33.86, d.f. = 4, P < 0.001), indicating that there
were differences among populations in larval performance
Fig. 6. Relationship between mean adult Y. mahalebella body mass
and mean temperature recorded at different populations during the
larval development period. Symbols represent mean values, and
bars = ±SEM. Populations are ordered by altitude from lowest (Poyo
Manquillo) to highest (Cabeza del Tejo) elevation.