Do you want to buy antibiotics online without prescription? https://buyantibiotics24h.net/ - This is pharmacy online for you!
Angiotensin-converting enzyme-like activity in crab gills and its putative role in degradation of crustacean hyperglycemic hormone
Archives of Insect Biochemistry and Physiology 68:171–180 (2008)
Angiotensin-Converting Enzyme-Like Activity in CrabGills and Its Putative Role in Degradation ofCrustacean Hyperglycemic Hormone
J. Sook Chung1* and S.G. Webster2
Angiotensin-converting enzyme-like enzyme activity (ACELA) was found in Carcinus maenas using reverse phase high perfor-mance liquid chromatography (RP-HPLC) analysis of degradation kinetics of a synthetic substrate (Hippuryl-histidyl-leucine)and a specific inhibitor (captopril). Gills contained the highest ACELA, then brain, muscle, and testis, respectively, while noactivity was detected in the following tissues: hepatopancreas, hindgut, hypodermis, heart, and hemolymph. ACELA present ingill membranes exhibited a Km of 0.23 mM and Vmax of 7.6 nmol with synthetic substrate. The enzyme activity was dependenton Cl– concentration and was markedly inhibited by captopril, lisinopril, and EDTA. Addition of Zn2+ to membranes previouslytreated with EDTA restored 89% activity, suggesting that C. maenas ACELA is a Zn2+ metalloenzyme. Gill membranes pre-pared from premolt crabs showed similar levels of ACELA to those of the intermolt animals. Administration of captopril in vivolengthened the half life of circulating CHH, while in vitro incubation of gill membranes with captopril reduced CHH. Theseresults suggest that C. maenas ACELA present in gills is likely to be involved in degradation of this neuropeptide. Arch. InsectBiochem. Physiol. 68:171–180, 2008.
KEYWORDS: angiotensin-converting enzyme-like activity; crab gills; crustacean hyperglycemichormone; inactivation of hormone
INTRODUCTION
iting hormone (MIH), and crustacean cardioactivepeptide (CCAP) in hemolymph are low, ranging in
concentration from <10–11 to 10–10 M (Webster, 1996;
lymph or blood are transient and at low concentra-
Chang et al., 1998; Chung et al., 1999; Webster and
tion, yet they are crucial for initiating signalling
Chung, 1999; Phlippen et al., 2000; Chung and
pathway resulting in physiological action. In crus-
Webster, 2005; Nakatsuji and Sonobe, 2003).
taceans, crustacean hyperglycemic hormone (CHH)
Compared with crustacean neuropeptides (5–
is usually released in immediate response to stresses
10 min), insect neuropeptides, despite relatively
(e.g., hyperthermal, hypoxia, and emersion), but
smaller size, appear to have much longer half lives
stays only a short time in the hemolymph (Webster,
in circulation. For example, adipokinetic hormone,
1996; Chang et al., 1998; Chung and Webster,
hypertrehalosemic hormone, allatostatins, and
2005). The circulating levels of CHHs, molt inhib-
bombyxin, when injected into various species of
1Center of Marine Biotechnology, University of Maryland Biotechnology Institute, Baltimore, Maryland2School of Biological Sciences, Bangor University, Bangor, Gwynedd, United Kingdom
Grant sponsor: Biotechnology and Biological Sciences Research Council, UK.
Abbreviations used: ACE = angiotensin converting enzyme (dipeptidyl carboxypeptidase I, EC 3.4.15.1); AG-I = angiotensin-I; CHH = crustaceanhyperglycemic hormone.
*Correspondence to: J. Sook Chung, Center of Marine Biotechnology, University of Maryland Biotechnology Institute, Baltimore, MD 21202. E-mail: chung@comb.umbi.umd.edu
Received 1 December 2007; Accepted 4 March 2008
2008 Wiley-Liss, Inc. DOI: 10.1002/arch.20247Published online in Wiley InterScience (www.interscience.wiley.com)
insects, showed half lives of approximately 18–
Here we report that ACELA is, indeed, present
53 min (Rayne and O’Shea, 1992; Strey et al., 1993;
in gills of C. maenas. The results obtained from in
Garside et al., 1997; Goldsworthy et al., 2002) and
vitro incubation of CHH with gill membranes and
up to 170 min (Suenobu et al., 2004) in hemo-
an in vivo half life study suggest that ACELA may
lymph. Such short half lives of crustacean neuro-
be an enzyme involved in regulation of CHH titres
peptides, compared to insects, imply that crustaceans
in hemolymph, and CHH can be an endogenous
may have mechanisms to rapidly remove circulat-
ing neuropeptides. However, very little is knownabout enzymes and tissues in crustaceans that
MATERIALS AND METHODS
may be involved in processes of metabolic inac-
tivation, degradation, or in clearance of circu-lating neuropeptides. C. maenas were collected with baited traps
Much of the information available in inverte-
from the Menai Strait (Wales, UK). They were kept
brates regarding the degradation of neuropeptides
in re-circulated seawater at 12–15°C under ambi-
or neurohormones was initially gathered through
ent light conditions. Adult male crabs at intermolt
homologous studies of vertebrate systems. For ex-
(carapace width: 45–55 mm) were used for all
ample, angiotensin-converting enzyme (ACE)
activity in invertebrates has been suggested to inac-tivate neuropeptides (Lamango and Issac, 1994;
Chemicals
Laurent and Salzet, 1996a,b; Lamango et al., 1996). Moreover, the wide distribution of ACE activity, in-
Hippuryl-1-histidyl-1-leucine (Hip-His-Leu),
cluding central nervous tissues, reproductive tissues,
hippuric acid, captopril {1-[(2S)-3-mercapto-2
hemocytes, and hemolymph, may further implicate
methylproprionyl]-L-proline}, and lisinopril {(S)-
ACE in a putative role in other physiological actions
1{N2-(1-carboxy-3-phenylpropyl)-L-lysyl]-L-pro-
such as metamorphosis, reproduction, and immu-
line dihydrate} were purchased from Sigma (St.
nity in corresponding tissues (Vandenbroeke et al.,
Louis, MO). Angiotensin I was obtained from
1997; Hens et al., 2002; Vandingenen et al., 2002;
Ekbote et al., 2003; Salzet and Verger-Bocquet, 2001;Macours et al., 2003; Laurent et al., 1997; Leung et
Purification, Quantification,
al., 1992). In crustaceans, angiotensin-converting
and Iodination of CHH
enzyme-like activity (ACELA) was initially reportedin the gills of the blue crab, Callinectes sapidus
The methods for isolation, purification, and
(Smiley and Doig, 1994) and in the terrestrial crab,
quantification of C. maenas CHH and iodination
Chasmagnathus granulatus (Delorenzi et al., 1996).
procedures were as described in Webster (1993).
Our initial clearance experiment to determine
a half-life of CHH using [125I]CHH (previously in-
Briefly, the extracts of sinus glands were first iso-
jected into crabs) showed that it was rapidly re-
lated on a Phenyl column (4.6 × 250 mm, Wa-
moved from hemolymph, and accumulated in the
ters) using a gradient 30–80% B over 60 min at a
gills. This result indicated that gills may play a role
flow rate of 1.0 ml/ min. (A = 0.11% trifluoroacetic
in degradation of CHH. Our preliminary observa-
acid [TFA] in H2O; B = 0.1% TFA in 60% acetoni-
tion, together with previous reports that ACE ac-
trile.) The fractions containing CHH were then
tivity was found in gills of other species of crabs
pooled and re-run on a C18 column (4.6 × 250
(Smiley and Doig, 1994; Delorenzi et al., 1996),
mm, Phenomenex) using a gradient 45–60% B
led us to investigate ACELA in C. maenas gills and
over 30 min at same flow rate. Peaks were manu-
its putative role in regulating neuropeptide levels
ally collected. The quantification of neuropeptides
was performed by amino acid analysis of hydroly-
Archives of Insect Biochemistry and Physiology
sates obtained by gas-phase hydrolysis in vacuo
gently overlaid on 200 µl of the same buffer con-
at 150°C for 1 h using azeotropic hydrochloric
taining 0.06% (w/v) Triton X-114 and 6% (w/v)
acid containing a trace of phenol. Vacuum-dried
sucrose and incubated for 5 min at 37°C. Phase
hyrolysates were then quantified for amino acid
separation of Triton X-114 was obtained by imme-
composition by o-phthalaldehyde pre-column
diate centrifugation at 13,000 rpm for 5 min at
derivertization. Then, 300 pmol of CHH was iodi-
20°C. The supernatant was collected and the de-
nated with 300 µCi of [125I](Amersham) using the
tergent layer was re-suspended in an equal volume
of buffer (50 mM Tris/HCl, 150 mM NaCl, pH7.4). Both detergent-rich and detergent-poor frac-
Preparation of Membranes
Anterior and posterior gills were dissected from
ACE Assay
crabs (n = 6), after being pre-chilled on ice for 30min. After rinsing in ice-cold crustacean saline
(Webster and Keller, 1986), gills were homogenized
brane preparations (20 µg/20 µl final incubation
with a Polytron® in ice-cold buffer containing 50
volume) in the presence of 3 nmol of Hip-His-
mM HEPES, either in the presence or absence of
Leu in the assay buffer described above, either in
300 mM NaCl (pH 8.3). Homogenates were cen-
the presence or absence of captopril (final concen-
trifuged at 1,000g for 10 min at 4°C and superna-
tration at 18.75 µM) for 30 min at 37°C. The en-
tants were further centrifuged at 30,000g for 40 min
zyme activity was calculated by the estimation of
at 4°C. The resulting pellets, once re-suspended in
the difference between amounts of remaining Hip-
50 mM HEPES, were estimated for protein con-
His-Leu at times 0 and 30 min after incubation.
centration using a modified Lowry method (Lowry
At the beginning or end of incubation, samples
et al., 1951). Aliquots of membrane protein (120–
were acidified with glacial acetic acid (to give a
150 µg/µl) were immediately frozen in liquid ni-
final concentration of 2 N in 100 µl final volume).
trogen and kept at –20°C until used.
The samples were mixed and centrifuged at 13,000rpm for 5 min at room temperature. The superna-
Treatment of Gill Membranes With
tant (~100 µl) was applied onto a RP-HPLC con-
Ethyldiaminetetraacetic Acid (EDTA)
nected to a C18 column (Bakerbond WP C18, 4.6× 250 mm, J.T. Baker Co.), with a gradient of 30–
Membranes, pre-incubated with 5 mM EDTA in
80% B over 30 min (A = 0.11% TFA in H2O, B =
the assay buffer (50 mM HEPES and 300 mM NaCl,
0.1% TFA in 60% acetonitrile). Absorbance was
pH 8.3) for 30 min at 37°C, were centrifuged at
monitored at 210 nm with a flow rate of 1 ml/ min.
30,000g for 40 min at 4°C. Pellets were washed
The samples of AG-I and CHH were chromato-
twice with the assay buffer and the final pellets
were re-suspended in the same buffer. Degradation of Angiotensin-I and CHH Phase Separation of Membrane Proteins by C. maenas ACELA in Gills and With Triton X-114 Amino Acid Analysis of Peaks
Membranes were initially incubated on ice (30
Substrate specificity of C. maenas ACELA in gills
min with occasional shaking) with 10 mM Tris
was tested using synthetic ACE substrate, AG I, and
buffer (pH 7.4) containing 150 mM NaCl and
CHH. Each substrate (3 nmol) except CHH (300
1.5% (w/v) Triton X-114. Detergent-insoluble ma-
pmol) was incubated with a membrane concen-
terial was removed by centrifugation at 30,000g for
tration of 40 µg/50 µl at 37°C for 30 min, either
40 min at 4°C. Portions of the supernatant were
in the presence or in the absence of captopril (360
Archives of Insect Biochemistry and Physiology
µM). The samples were treated as described above
Gill membranes prepared from premolt crabs
for RP-HPLC separation. The degradation rate of
showed similar levels of enzyme activity (0.995 ±
each substrate was calculated by comparing peak
0.025 nmol/min/mg protein) to that of intermolt
areas of substrate in the presence or in the absence
(1.044 ± 0.05 nmol/min/mg protein).
of captopril. Peaks were collected manually andanalyzed for amino acid composition as described
Characteristics of ACELA in Crab Gills
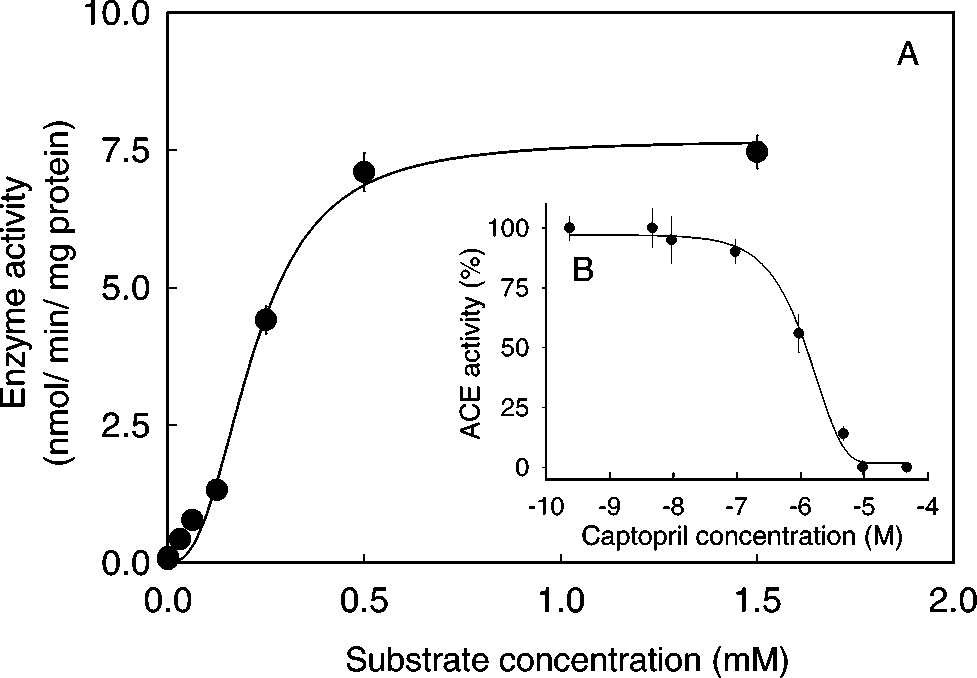
The saturation curve analyses of Lineweaver-
Statistics
Burk plot (Fig. 2) of the hydrolysis of Hip-His-Leuby ACELA in gill preparation gave values of Km,
Statistical analysis was carried out using either
0.23 ± 0.012 mM and Vmax, 7.6 nmol/min/mg
Student’s t-test or ANOVA (GraphPad InStat ver-
protein, respectively. All substrate concentrations
sion 3.0, GraphPad Software, San Diego, CA).
used were well below the Km value and in no casewas more than 10% of the substrate consumed dur-
ing the experimental period. Captopril inhibited
The Tissue Distribution of ACELA in Crabs
ACE activity in a dose-dependent manner with avalue of IC50 < 1 µM (Fig. 2B).
The following tissues from male intermolt C.
To determine the effect of Cl– concentration on
maenas were examined for ACELA: muscle (from
the hydrolysis of ACE substrate by gill membrane
walking legs), hypodermis, hindgut, heart, midget
preparations, NaCl was added to the incubation
gland, brain, hemolymph, and gills. Of these tis-
medium to give final concentrations of Cl– rang-
sues tested, gills displayed by far the highest level
ing from 0 to 300 mM. ACELA was highly depen-
of ACELA (Fig. 1). Brains, and to a lesser extent
dent on Cl– concentrations. The maximal activity
testis and muscle membranes, showed much less
was observed at 25 mM of Cl–, double the activity
enzyme activity than that of gill membranes (Fig.
compared to that at 0 mM (Fig. 3). Increasing Cl–
1). Gill membranes prepared from the edible crab,
concentration to over 300 mM did not change the
Cancer pagurus, also exhibited ACELA, with approxi-
mately 30% of that in C. maenas gills. A: Michaelis-Menten plot of the hydrolysis of
Tissue distribution of ACELA. Enzyme activity
synthetic ACE substrate by Carcinus gill membranes. B:
(pmol/min/mg protein) was presented as mean ± 1 SE, n
ACELA was inhibited by captopril in a dose-dependent
= 5–8. ND = non-detected. Statistical significance was
manner. Each point in A and B was shown as mean ± 1
tested using nonparametric ANOVA (Kruskal-Wallis Test).
Archives of Insect Biochemistry and Physiology
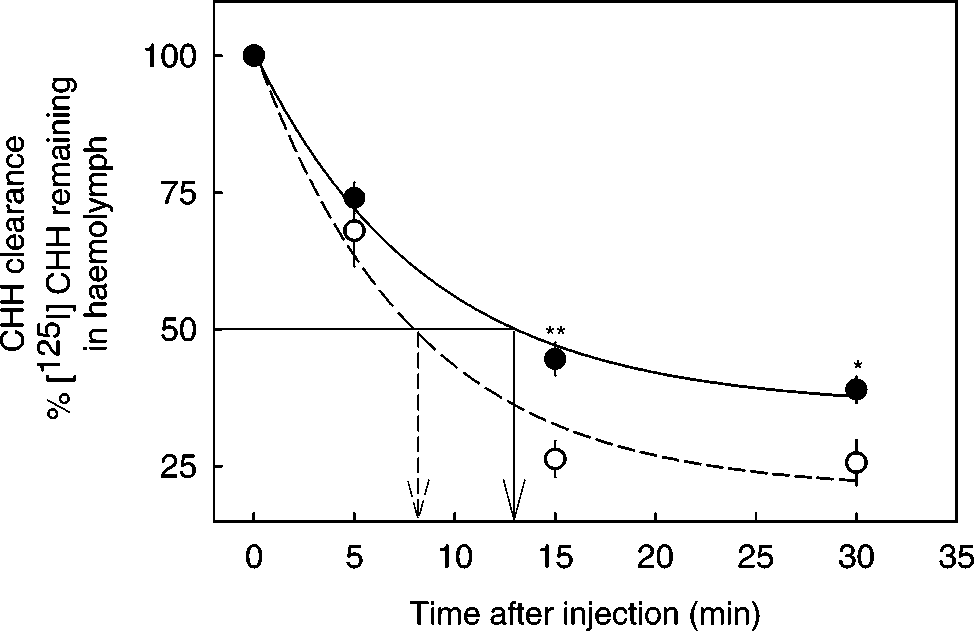
In Vivo Clearance Rate of CHH
Initially, 120 fmol of [125I] CHH (~200,000
DPM) was injected into crabs. Two minutes afterinjection (to allow distribution of the injected bo-lus), the radioactivity in the hemolymph was mea-sured and was considered to be at a reference time0 (100%). At 5, 10, and 30 min, small hemolymphsamples (100 µl) were removed and radioactivitywas measured by a gamma counter (Wallac 1470). The hormone was cleared very rapidly, with an es-timated half-life of 8 min (Fig. 4). When crabs werefirst injected with captopril, to give an estimated
Effect of Cl– concentrations on ACELA in gills.
circulating concentration of 500 µM, followed by
Each point was presented as mean ± 1 SE (n = 4–6).
hormone injection, the half-life increased to 13min. At sampling points of 15 and 30 min, crabs
Effect of a Divalent Chelating Agent (EDTA) and
injected with captopril showed a significantly high
Divalent Metal Ions on the ACELA
recovery of [125I]CHH in their hemolymphs com-pared to controls (P > 0.01 and P > 0.05, respec-
EDTA (100 µM) -treated gill membranes con-
tively). Subsequent dissection of major tissues from
tained 21% of enzyme activity compared with un-
these crabs (gills, hindgut, hepatopancreas, testis,
treated controls. Inclusion of the following bivalent
heart, muscle, and hypodermis) followed by the
metal ions (Co2+, Cu2+, Ca2+, Mg2+, Mn2+, Fe2+, andZn2+) was carried out to determine whether enzymeactivity could subsequently be retrieved from thegill membranes pre-treated with 5 mM of EDTA. Theaddition of 100 µM of either Zn2+ or Co2+ restoredapproximately 89 and 56% of enzyme activity, whileMn2+, Mg2+, or Fe2+ application regained less activ-ity (37, 19, and 13%, respectively). In contrast, theremaining ACELA in the membrane pre-treatedwith EDTA was completely abolished by the addi-tion of 100 µM of Cu+. Phase Separation of C. maenas ACELA
The phase separation property of Triton X-114
The clearance of [125I] CHH from hemolymph,
at >30°C in partitioning hydrophobic proteins into
in vivo. Animals were injected with [125I]CHH and
the detergent-rich phase has been used for isolat-
hemolymphs were removed at the sampling points. Thesamples were counted for the radioactivity remaining in
ing integral membrane proteins. ACELA was esti-
hemolymphs. Closed circles: In the absence of captopril;
mated after gill membrane was treated with Triton
Open circles: in the presence of captopril. Each point was
X-114, to determine if this enzyme activity was
shown as mean ± 1 SE (n = 8). Statistical significance was
associated with an integral membrane protein.
obtained by the comparison between in the absence and
Approximately 80% of the activity was in the de-
the presence of captopril at sampling points of 15 min
tergent-poor phase, while the remaining activity
(**) and 30 min (*), respectively. **P < 0.005, *P < 0.05
(20%) was recovered from the detergent-rich phase.
Archives of Insect Biochemistry and Physiology
measurement of residual radioactivity revealed thatalmost 98% of the radioactivity was found in gills. The radioactivity was equally distributed over gillpairs, and there was no difference between ante-rior and posterior gills (Fig. 5). Degradation of Angiotensin-I and CHH by C. maenas ACELA in Gills
To determine substrate specificity of ACELA in
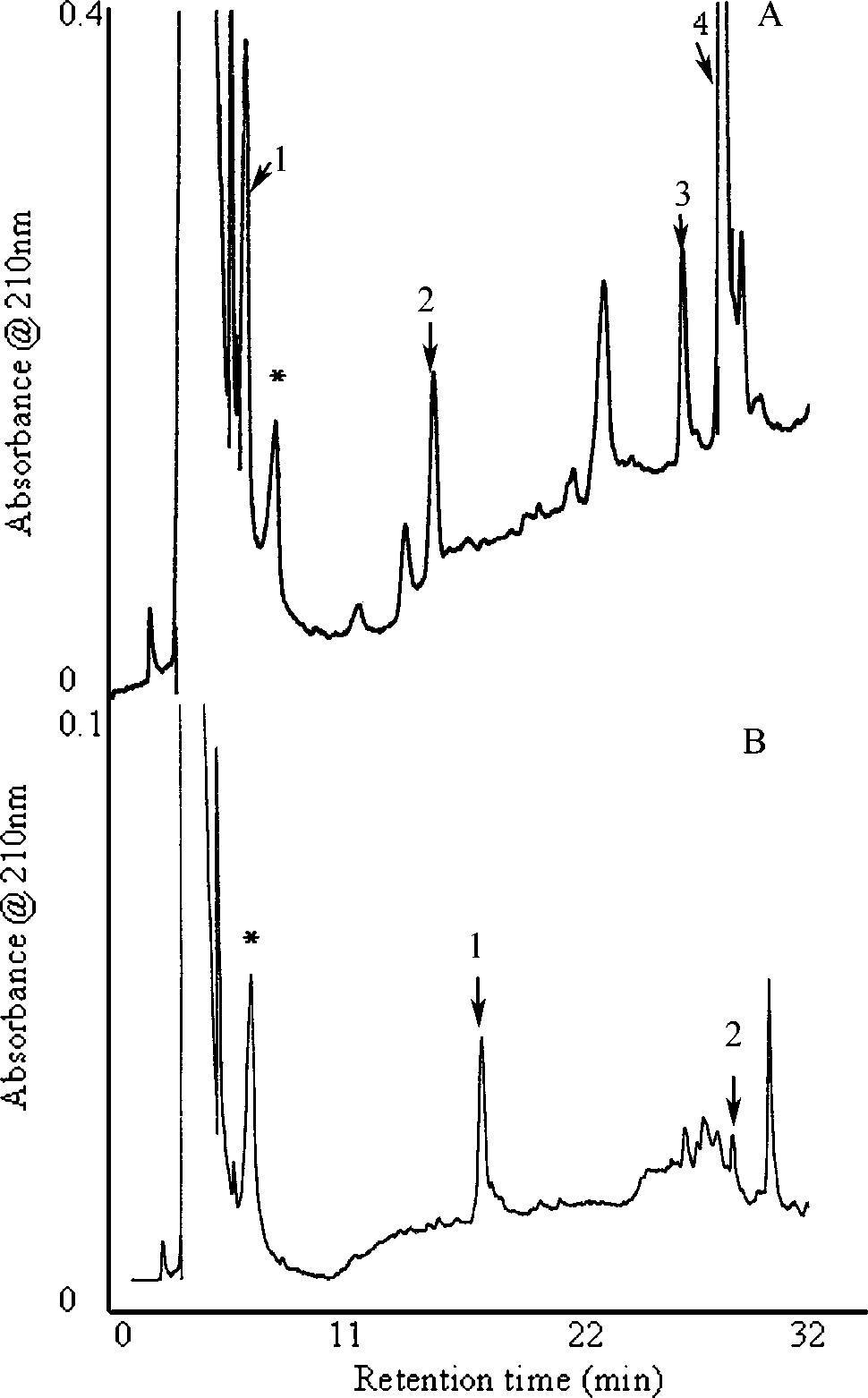
C. maenas gills, Hip-His-Leu, AG-I, and CHH wereincubated with gill membranes in the presence ofcaptopril. The presence of captopril at a concen-tration of 360 µM reduced the degradations of Hip-His-Leu by 46%, of AG-I, by 13%, and of CHH by83%. Peaks identified by amino acid analyses showin Figure 6A that C. maenas ACELA processed Ag-I(peak 4) to Ag-II (peak 3) by cleaving off His-Leu-OH (peak 1) from the C-terminus. Peak 2 (AG-III), the product of peak 3, was further cleaved byaminopeptidase. Amino acid composition of peak1 of Figure 6B (Table 1) showed that it was a de-graded product of CHH, while peak 2 was identi-fied as intact CHH.
RP-HPLC chromatogram showed cleavage of AG-
I (A) and CHH (B) after incubation with membranes pre- pared from gills. The results of amino acid composition of each peak are as follows. A: Peak 1: His-Leu-OH; Peak 2 (AG-III): Arg-Val-Tyr-Ile-His-Pro-Phe-OH; Peak 3 (AG- II): Asp-Arg-Val-Tyr-Ile-His-Pro-Phe-OH; and Peak 4: H- Asp-Arg-Val-Tyr-Ile-His-Pro-Phe-His-Leu-OH. B: Peak 1: Digested CHH; Peak 2: CHH intact. *Captopril. Note that the peaks obtained from each HPLC do not accurately represent amount of product formed due to a difference in extinction coefficients.
The distribution of injected [125I] CHH in gills. DISCUSSION
Gills were collected 30 min after the injection of radioac-tive CHH. After rinsing and drying, the radioactivity was
The present study using the synthetic substrate
measured. Anterior gills (4) were presented as A1–A4; pos-terior gills, P1–P4. Bar was presented as mean
of mammalian ACE (Hip-His-Leu-OH) and cap-
8). There was no statistical significance (at P < 0.05)
topril, a selective specific inhibitor, demonstrated
among the values of [125I] CHH obtained from gills (non-
ACELA in gill, brains, and testis of the shore crab,
parametric ANOVA, Kruskal-Wallis Test). C. maenas. The highest ACELA was detected in the
Archives of Insect Biochemistry and Physiology
TABLE 1. The Amino Acid Composition of Each Peak (Fig. 6A and B)*
1997). However, ACELA was not detected in C.maenas hemolymph, while similar activity was de-
scribed in hemolymph of Mytilus edulis (Leung et
By injection of [125I] CHH into C. maenas, it
was possible to monitor the clearance rate from
hemolymph as well as tissue-specific accumulation
of this radiolabelled peptide. The measured half-
life of C. maenas [125I] CHH in hemolymph was
5–10 min, in agreement with our previous results
obtained from non-radiolabelled CHH injection
into crabs (Webster, 1996; Chung and Webster,
1996). However, in the presence of captopril, the
half-life of this peptide was increased by more than
*Amino acids (AA) are presented as three letter abbreviation. Numbers shown in
5 min. Over 99% of the radioactivity was recov-
upper layer were obtained from the amino acid analysis of the peaks shown inFigure 6A and B, while those in parentheses are based on the amino acid se-
ered from the gill tissues, in which it was distrib-
quences of peptides and their fragments. a
uted equally between the left and right sides of
the gills. Posterior gills showed slightly higher ra-dioactivity than those of anterior gills, but the dif-
gill membranes. Enzyme activities in gills of pre-
ference was not statistically different. This implies
molt crabs were similar to activities of intermolt
that the distribution of ACELA is ubiquitous,
animals, suggesting that this enzyme is probably
throughout posterior and anterior gills.
not involved in the molting process. The wide dis-
C. maenas ACELA in gills produced AG-II by
tribution of ACELA in various tissues of C. maenas
cleaving the dipeptide from the C terminus of AG-
is in agreement with previous results found in the
I (Fig. 6A). Interestingly, AG-II was further pro-
blue crab, C. sapidus (Smiley and Doig, 1994). In-
cessed by an aminopeptidase that was co-present
terestingly in C. granulatus, ACELA found in CNS
in gill membranes, yielding a further product,
and gills was co-exited with AGII (Delorenzi et al.,
peak 2 (AG-III). The combination of selective in-
1996).Additionally, the gill membranes of the ed-
hibitors of neprilysin (phosphoramidon and 1,10
ible crab, C. pagurus, also exhibited ACELA, sug-
o-phenanthroline) and aminopeptidase (ama-
gesting that ACELA in gills may be a common
statin) with these substrates showed that each of
these peptides can be degraded by more than one
Based on our results, the properties of C. maenas
enzyme in the tissue membrane extract (Masler
ACELA were also similar to those of mammalians
et al., 1996; Leung et al., 1992). Surely, synthetic
and invertebrates including houseflies and leeches
ACE substrate containing a single ACE-specific
(Salzet and Verger-Bocquet, 2001, Ekbote et al.,
cleavage site can be the only substrate for ACE,
2003; Macours et al., 2003). The identical proper-
whereas AG-I might be a substrate for neprilysin
ties are listed as follows: (1) Zn2+ metalloenzyme,
in the presence of captopril. As a result, the in-
(2) captopril and lisinopril sensitive, and (3) Cl–-
hibitory effect of captopril on AG-I degradation
dependent activity. Furthermore, C. maenas ACELA
was much smaller than that of the synthetic sub-
seemed to be a membrane-bound protein, but it
strate. This result is congruent with a finding from
could be easily dissociated from the membrane.
the leech (Ekbote et al., 2003), also suggesting
After phase separation with Triton X-114, most of
the presence of multiple enzymes in the gill mem-
the enzyme activity was found in the detergent-
poor phase, like the one in the leech and housefly
The highest inhibition (83%) of ACE in the pres-
(Salzet and Verger-Bocquet, 2001; Laurent et al.,
ence of captopril was observed with C. maenas CHH
Archives of Insect Biochemistry and Physiology
pathway of AG-I (A) and pu- tative cleavage sites of ACELA in CHH (B). Amino acids itali- cized: cleavage sites; amino ac- ids underlined: parts possibly cleaved.
compared with AG-I and synthetic ACE substrate.
in gills agreed with a broad range of enzyme activ-
Considering the nature of known ACE substrates,
ity including endopeptidase function on a wide
CHH would seem an unlikely candidate since it is
substrate specificity (Isaac et al., 1998).
large (72 amino acid residues), blocked at both
Crustacean gills, which are multifunctional tis-
N- and C-termini, and has three intradisulphide
sues composed of at least six different cell types
bridges (Kegel et al., 1989). The solution structures
(Mantel and Farmer, 1983; Lawson et al., 1994),
of CHH neuropeptides (CHH and MIH) obtained
seem to serve as a target tissue of the CHH recep-
by circular dichroism consisted of a very high per-
tor (Chung and Webster, 2006) as well as a site of
centage of α-helices and β-strands (Chung and
degradation. It will be of interest to localize the
Webster, 1996; Katayama et al., 2003). A close ex-
amination of the primary sequence of CHH reveals
In conclusion, we propose that ACELA present
that there are more than five putative ACE cleav-
in the gill membranes of C. maenas participates in
age sites (Fig. 7B), but these putative cleavage sites
regulating the levels of circulating neuropeptides.
may not be readily available to ACE since they may
Additionally, since ACELA is found in the gills of
not be exposed to the surface of the molecule. Nev-
following species, C. maenas, C. pagurus, C. sapidus,
ertheless, our preliminary data obtained from the
and C. granulatus, we believe that this enzyme may
amino acid analysis of two peaks of CHH cleaved
be ubiquitously present in all crustaceans with at
by ACELA indicated that the cleavage occurred at
least a function in the processing/inactivation/deg-
a part of the C-terminus and potentially the area
radation of circulating neuropeptides.
between disulphide bridges, resulting in frag-mented and inactivated CHH. However, we are un-
ACKNOWLEDGMENTS
able to locate the exact cleavage site of CHH. Morework is required to obtain detailed information on
This work was supported by a grant to S.G.W.
the degradation pathway of CHH. Our initial study
from the Biotechnology and Biological Sciences
into the substrate specificity of crab ACELA present
Archives of Insect Biochemistry and Physiology
LITERATURE CITED
advances in arthropod endocrinology, Society for Experi-
mental Biology Seminar Series. Cambridge: Cambridge
Chang ES, Keller R, Chang SA. 1998. Quantification of crus-
tacean hyperlgycemic hormone by ELISA in hemolymph
Katayama H, Nagata K, Ohira T, Yumoto F, Tanokura M,
of the lobster, Homarus americanus, following various
Nagasawa H. 2003. The solution structure of molt-inhib-
stresses. Gen Comp Endocrinol 111:359–366.
iting hormone from the Kumara prawn Marsupenaeus
Chung JS, Webster SG. 1996. Does the N-terminal pyro-
japonicus. J Biol Chem 278:9620–9623.
glutamate residue have any physiological significance for
Kegel G, Reichwein B, Weese S, Gaus G, Peter-Katalinic J,
crab hyperglycaemic neuropeptides? Eur J Biochem 240:
Keller R. 1989. Amino acid sequence of the crustacean hy-
perglycemic hormone (CHH) from the shore crab, Carcinus
Chung JS, Webster SG. 2005. Dynamics of in vivo release of
maenas. FEBS Lett 255:10–14.
molt-inhibiting hormone (CHH) and crustacean hyper-
Lamango NS, Isaac RE. 1994. Identification and properties
glycaemic hormone (CHH) in the shore crab, Carcinus
of a peptidyl dipeptidase in the house fly, Musca domestica,
maenas. Endocrinology 146:5545–5551.
that resembles mammalian angiotensin-converting en-
Chung JS, Dircksen H, Webster SG. 1999. A remarkable, pre-
cisely timed release of hyperglycemic hormone from endo-
Lamango NS, Sajid M, Isaac RE. 1996. The endopeptidase
crine cells in the gut is associated with ecdysis in the crab
activity and the activation by Cl– of angiotensin-convert-
Carcinus maenas. Proc Natl Acad Sci USA 96:13103–13107.
ing enzyme is evolutionarily conserved: purification and
Delorenzi A, Pedreira ME, Romano A, Garcia SY, Pirola CJ,
properties of an angiotensin-converting enzyme from the
Nahmod VE, Maldonado H. 1996. Angiotensin II enhances
housefly, Musca domestica. Biochem J 314:639–646.
long-term memory in the crab Chasmagnathus granulatus.
Laurent V, Salzet M. 1996a. Biochemical properties of the
angiotensin-converting like enzyme from the leech Thero-
Ekbote UV, Weaver RJ, Isaac RE. 2003. Angiotensin I- con-
myzon tessulatum. Peptides 17:737–745.
verting enzyme (ACE) activity of the tomato moth,
Laurent V, Salzet M. 1996b. Metabolism of angiotensins by
Lacanobia oleracea: changes in levels of activity during de-
head membranes of the leech Theromyzon tessulatum. FEBS
velopment and after copulation suggest roles during meta-
morphosis and reproduction. Insect BiochemMol Biol
Laurent V, Stefano G, Salzet M. 1997. Presence and biochemi-
cal properties of a molluscan invertebrate angiotensin-con-
Garside CS, Hayes TK, Tobe SS. 1997. Inactivation of Dip-
verting enzyme. Regul Pept 69:53–61.
allatostain 5 by membrane preparations from the cock-
roach Diploptera punctata. Gen Comp Endocrinol 108:
Lawson SL, Jones MB, Moate RM. 1994. Structural variabil-
ity and distribution of cells in a posterior gill of Carcinusmaenas (Decapoda: Brachyura). J Mar Biol Ass UK
Goldsworthy GJ, Kodrik D, Comley R, Lightfoot M. 2002. A
quantitative study of adipokinetic hormone of the fire-
bug, Pyrrhocoris apterus. J Insect Physiol 48:1103–1109.
Leung MK, Houston B, Stefano GB. 1992. Degradation of Met-
enkephalin by hemolymph peptidases in Mytilus edulis.
Hens K, Vandingenen A, Macours N, Baggerman G, Karao-
glanovic AC, Schoofs L, De Loof A, Huybrechts R. 2002.
Characterization of four substrates emphasizes kinetic
Lowry OH, Rosebrough NJ, Farr AL, Randall RJ. 1951 Protein
similarity between insect and human C-domain angio-
measurement with the Folin phenol reagent. J Biol Chem
tensin-converting enzyme. Eur J Biochem 269:3522–3530.
Isaac RE, Coates D, Williams TA, Schoofs L. 1998. Insect an-
Macours N, Hens K, Francis C, De Loof A, Hybrechts R. 2003.
giotensin-converting enzyme: comparative biochemistry
Molecular evidence for the expression of angiotensin con-
and evolution. In: Coast GM, Webster SG, editors.Recent
verting enzyme in hemocytes of Locusta migratoria: stimu-
Archives of Insect Biochemistry and Physiology
lation by bacterial lipopolysaccharide challenge. J Insect
Strey A, Hayes TK, Isaac RE. 1993. Metabolism of insect
hypertrehalosemic hormone in Blaberus discoidalis cock-
Mantel LH, Farmer LL.1983. Osmotic and ionic regulation.
In Mantel LH, editor.The biology of crustacea: internal
Vandenbroeke F, Laurent V, Verger-Bocquet M, Stefano M,
anatomy and physiological regulation, vol. 5. New York:
Salzet M. 1997. Biochemical identification and ganglionic
localization of leech angiotensin-converting enzymes.
Brain Res Mol Brain Res 49:229–237.
Masler EP, Wagner RM, Kovaleva ES. 1996. In-vitro metabo-
lism of an insect neuropeptide by neural membrane prepa-
Vandingenen A, Hens J, Baggerman G, Macours N, Schoofs
rations from Lymantria dispar. Peptides17:321–326.
L, De Loof A, Huybrechts R. 2002. Isolation and charac-
terization of an angiotensin coverting enzyme substrates
Nakatsuji T, Sonobe H. 2003. Measurement of molt-inhibit-
from vitellogenic ovaries of Neobellieria bullata. Peptides
ing hormone titer in hemolymph of the American crayfish,
Procambarus clarkii, by time-resolved fluoroimmunoassay.
Webster SG. 1993. High-affinity binding of putative moult-
inhibiting hormone (MIH) and crustacean hyperglycaemic
Phlippen MK, Webster SG, Chung JS, Dircksen H. 2000.
hormone (CHH) to membrane bound receptors on the
Ecdysis of decapod crustaceans is associated with dramatic
Y-organ of the shore crab Carcinus maenas. Proc R Soc Lond
release of crustacean cardioactive peptide into the hemo-
Webster SG. 1996. Measurement of crustacean hyperglycaemic
Rayne RC, O’Shea M. 1992. Inactivation of neuropeptide hor-
hormone levels in the edible crab Cancer pagurus during
mones (AKH-I and AKH-II) studied in vivo and in vitro.
emersion stress. J Exp Biol 199:1579–1585.
Salzet M, Verger-Bocquet M. 2001. Elements of angiotensin
Webster SG, Chung JS. 1999. Roles of moult-inhibiting hor-
system are involved in leeches and molluscs immune re-
mone and crustacean hyperglycemic hormone in control-
sponse modulation. Brain Res Mol Brain Res 94:137–147.
ling moulting in decapod crustaceans. In: Roubos EW,
Bonga SEW, H Vaudry H, DeLoof A, editors. Recent de-
Smiley JM, Doig MT. 1994.Distribution and characterisation
velopments in comparative endocrinology and neurobi-
of angiotensin-converting enzyme-like activity in tissues
ology, Proceedings of the 19th Conference for European
of the blue crab, Callinectes sapidus. Comp Biochem Physiol
Comparative Endocrinology. Maastricht: Shaker. p 213–
Suenobu A, Mizoguchi A, Ichikawa T. 2004. Relationship be-
Webster SG, Keller R. 1986. Purification, characterization
tween firing activity of bombyxin-producing neurosecre-
and amino acid composition of the putative molt-inhib-
tory cells and hemolymph bombysin titer in the silkworm
iting hormone (MIH) of Carcinus maenas (Crustacea,
Bombyx mori. Gen Comp Endocrinol 137:219–226.
Decapoda). J Comp Physiol B 156:617–624.
Archives of Insect Biochemistry and Physiology
AMITY GLOBAL BUSINESS SCHOOL Module Syllabus Tourism is often claimed to be the biggest industry in the world, yet tourism is not really an industry, more a gathering together of disparate forms of production and consumptive activities. The distribution of this tourism is highly unequal geographically in terms of where tourists come from and where they travel to, though this is changing w
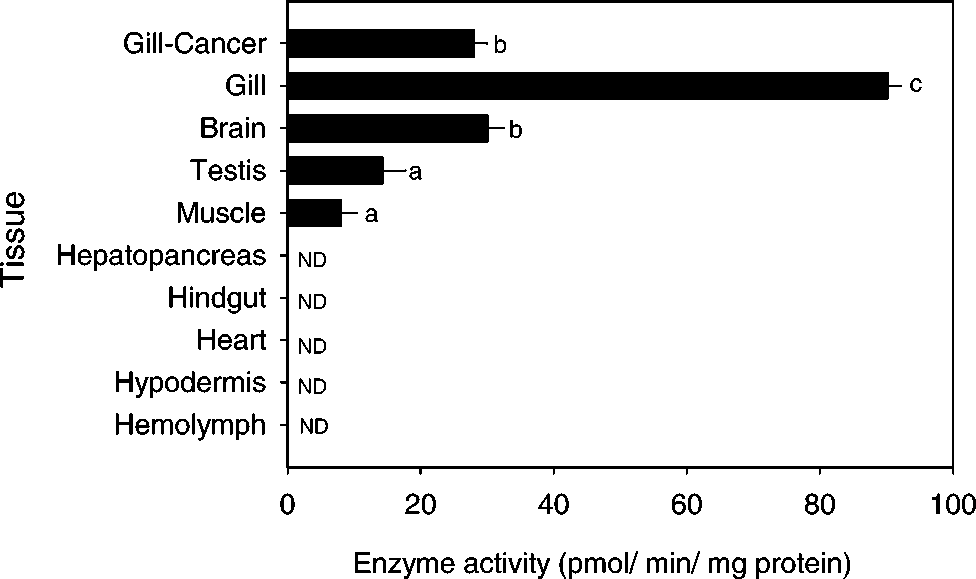
 µM). The samples were treated as described above
Gill membranes prepared from premolt crabs
for RP-HPLC separation. The degradation rate of
showed similar levels of enzyme activity (0.995 ±
each substrate was calculated by comparing peak
0.025 nmol/min/mg protein) to that of intermolt
areas of substrate in the presence or in the absence
(1.044 ± 0.05 nmol/min/mg protein).
µM). The samples were treated as described above
Gill membranes prepared from premolt crabs
for RP-HPLC separation. The degradation rate of
showed similar levels of enzyme activity (0.995 ±
each substrate was calculated by comparing peak
0.025 nmol/min/mg protein) to that of intermolt
areas of substrate in the presence or in the absence
(1.044 ± 0.05 nmol/min/mg protein).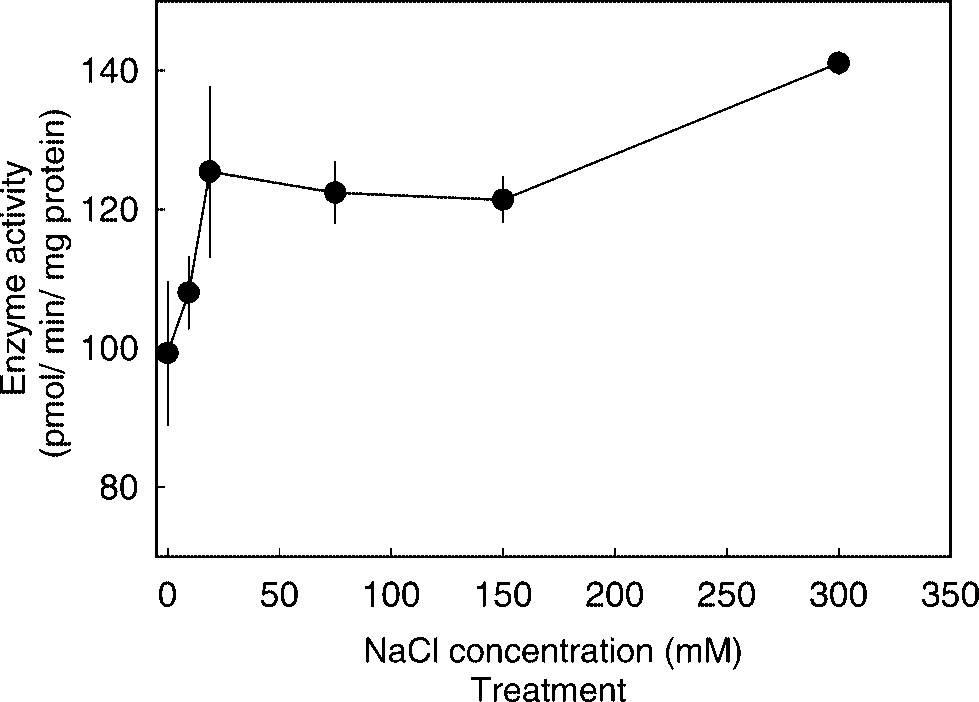
 In Vivo Clearance Rate of CHH
In Vivo Clearance Rate of CHH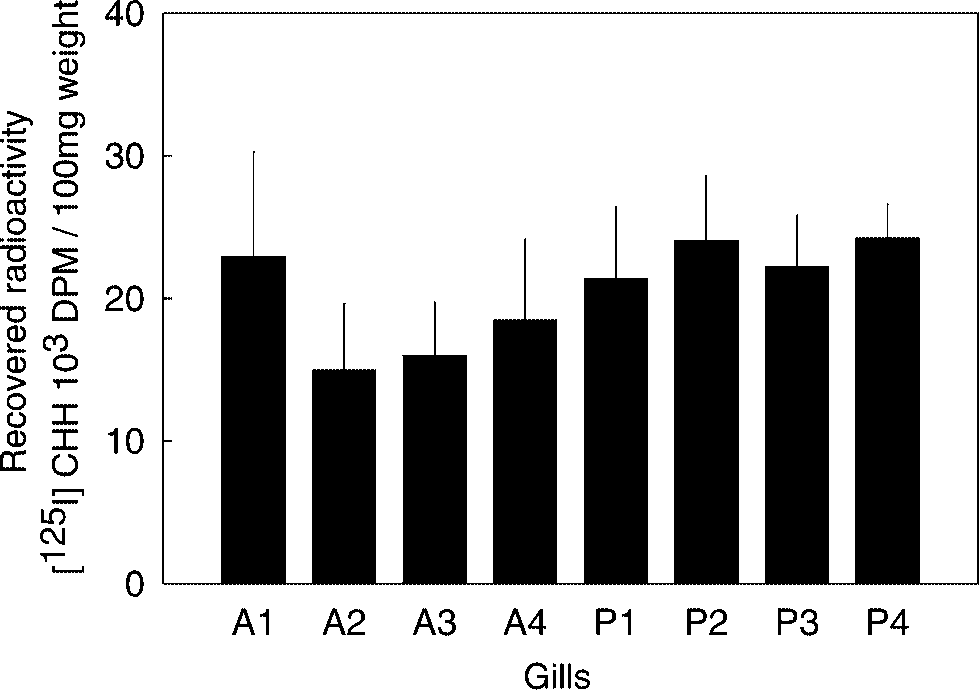
 measurement of residual radioactivity revealed thatalmost 98% of the radioactivity was found in gills.
measurement of residual radioactivity revealed thatalmost 98% of the radioactivity was found in gills.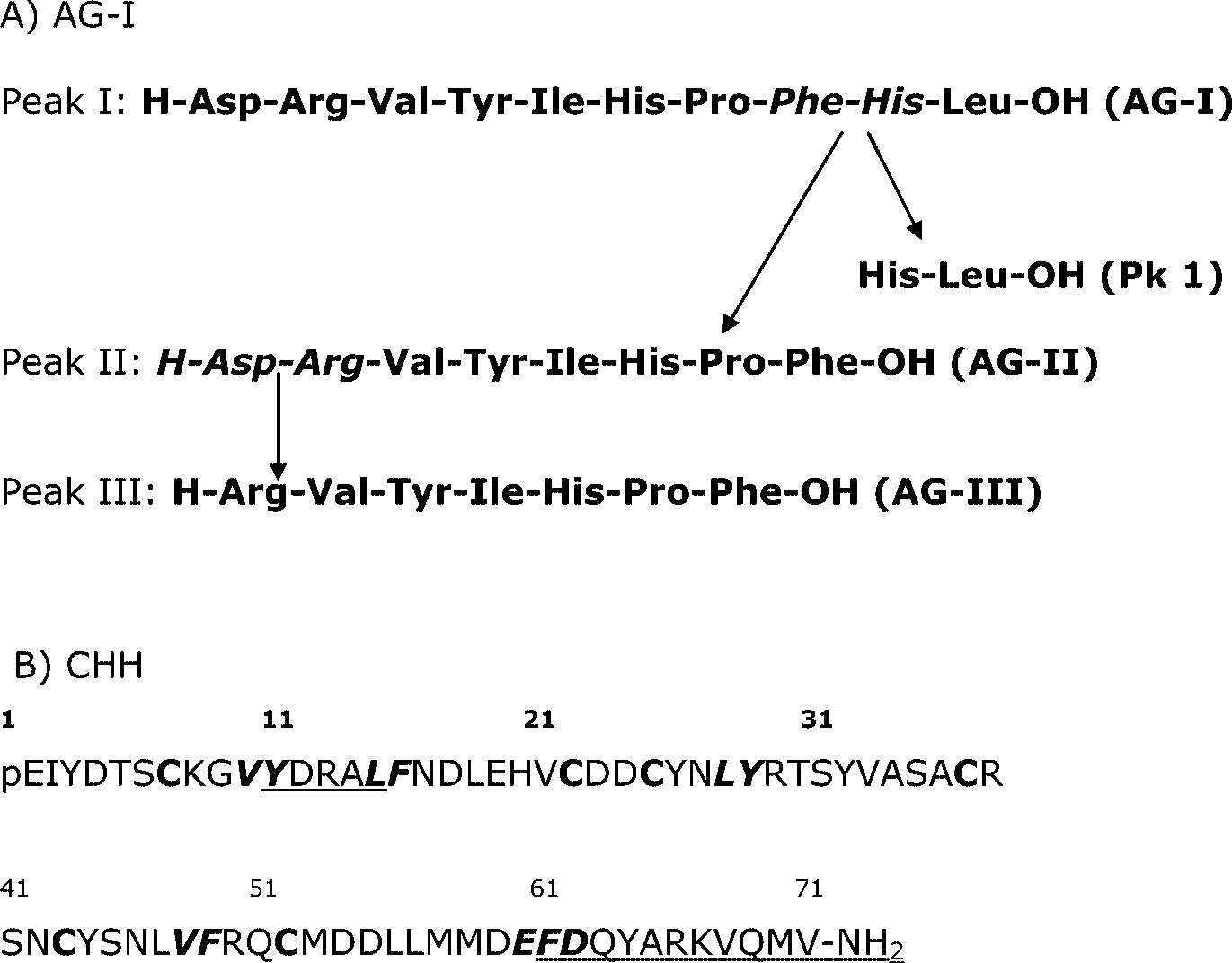
 pathway of AG-I (A) and pu-
pathway of AG-I (A) and pu-