Do you want to buy antibiotics online without prescription? https://buyantibiotics24h.net/ - This is pharmacy online for you!
Fungal transformation of an antimicrobial fluoroquinolone drug during growth on poultry litter materials
2004 Poultry Science Association, Inc. Fungal Transformation of an Antimicrobial Fluoroquinolone Drug During Growth on Poultry Litter Materials A. J. Williams,* I. A. Parshikov,† J. D. Moody,* T. M. Heinze,* and J. B. Sutherland*,1 *National Center for Toxicological Research, U. S. FDA, Jefferson, Arkansas 72079;and †Department of Medicinal Chemistry, University of Mississippi,Primary Audience: Researchers, Veterinarians, Public Health Officials The ability of a nonpathogenic fungus, Pestalotiopsis guepini, to metabolize fluoroquinolone antimicrobial agents during growth on poultry litter materials was investigated. Sterilized rice hulls, ground corncobs, and pine shavings in glass jars covered with foil were moistened with sterile water and inoculated with P. guepini. The litter materials then were dosed with norfloxacin and incubated for 20 d. In rice-hull cultures, P. guepini produced 4 metabolites: 7-amino-1-ethyl- 6-fluoro-4-oxo-1,4-dihydroquinolone-3-carboxylic acid, N-formylnorfloxacin, N-acetylnorfloxacin, and desethylene-N-acetylnorfloxacin. In corncob cultures, the fungus produced N-formylnorfloxacin and N-acetylnorfloxacin. In pine-shavings cultures, there was little growth of the fungus and no metabolism of norfloxacin. The results suggest that fungi that grow on poultry litter may degrade residues of antimicrobial drugs. Key words: fluoroquinolone, norfloxacin, poultry litter 2004 J. Appl. Poult. Res. 13:235–240 DESCRIPTION OF PROBLEM
that environment or are degraded by microor-ganisms that grow in the litter.
The use of antimicrobial agents in the poul-
Norfloxacin, a fluoroquinolone antimicro-
try industry to treat infections and promote
bial agent, is used clinically for the treatment
growth has frequently been associated with in-
of urinary tract infections, bacterial enteritis,and eye infections [6, 7, 8]. The same drug is
creases in bacterial resistance to clinically im-
also used in poultry production in some coun-
portant drugs [1, 2, 3, 4]. For instance, when
tries for chronic respiratory diseases caused by
fluoroquinolones are used, they may select for
Mycoplasma synoviae and Escherichia coli [9,
fluoroquinolone-resistant bacteria that can be
10, 11], although it is not registered for this
found in poultry litter [5]. It is not known,
however, whether all of the antimicrobial drugs
Poultry litter contains many bacteria, some
that reach poultry litter persist indefinitely in
of which may be resistant to multiple antibiot-
1 To whom correspondence should be addressed: jsutherland@nctr.fda.gov.
ics [5, 12, 13]. Although Enterococcus spp.,
Salmonella enterica, and other pathogenic bac-
equipped with an HP 1090L/M liquid chroma-
teria are sometimes found [13, 14, 15], they
tograph. A 2.0 × 250 mm Prodigy column was
usually are minor components of the total poul-
used with a water and acetonitrile gradient con-
try litter microbiota [16]. Saprobic fungi in the
taining 0.1% formic acid [21]. Full scans were
genera Aspergillus, Fusarium, Mucor, Penicil-
acquired in the positive-ion electrospray mode,
lium, and others are also found in poultry feed
with the capillary exit voltage at either +100
and litter [17, 18, 19, 20]. The objective of this
V or variable for molecular weight confirma-
study was to determine whether poultry litter
tion as the protonated molecule. The analysis
materials could affect the degradation of nor-
was repeated at +200 V to obtain fragments
floxacin by Pestalotiopsis guepini, a fungus
with [MH-H2O]+ as the base peak. Norfloxacin
known to metabolize fluoroquinolones [21].
metabolites were identified by comparing re-tention times and mass spectra with those pub-
MATERIALS AND METHODS
Typical poultry litter materials (10 g of rice
hulls, 10 g of pine shavings, or 20 g of ground
NMR) spectral analyses were performed at 500
corncobs) were placed in 500-mL mason jars
and sterilized by autoclaving for 1 h on each
[22] using deuterated methanol as the solvent.
of 2 successive days. Cultures of the fungus
The results were compared with those pub-
1325), grown on petri dishes of potato dextrose
RESULTS AND DISCUSSION
agar, were macerated in sterile water using ablender. Each jar of litter material was inocu-
Pestalotiopsis guepini grew well on rice
lated with 10 mL of the blended mycelium.
hulls and corncobs but not on pine shavings.
Norfloxacin was dissolved in 2% aqueous KOH
At 20 d, 4 metabolite peaks were detected by
(100 mg of norfloxacin per mL) and filter-steri-
HPLC in extracts from dosed rice-hull cultures
lized, and then 1 mL of this solution was added
that were not seen in extracts from control jars.
to each jar together with 40 mL of sterile water.
Two of the metabolite peaks were also detected
Controls were prepared using each of the types
in extracts from dosed corncob cultures. At 10
of litter material without the fungus or without
and 14 d, smaller concentrations of the metabo-
norfloxacin. The jars were incubated in the dark
lites were observed. Some additional peaks
at 28°C. On d 0, 10, 14, and 20, triplicate sets
were seen in the corncob and pine-shavings
of all cultures and controls were harvested,
cultures and controls; the mass spectra of these
filtered using glass wool, and extracted 3 times
peaks (not shown) indicated that they were not
with 100 mL of methylene chloride. The ex-
tracts were combined and then evaporated in
vacuo. The residues were dissolved in 2.0 mL
the peaks collected as they eluted from the
HPLC column were used to identify 4 norflox-
acin metabolites in the cultures of P. guepini
norfloxacin metabolites by high-performance
grown on rice hulls dosed with norfloxacin
liquid chromatography (HPLC), using a Hew-
(Figure 1A); the mass and NMR spectra were
lett-Packard Series 1100 liquid chromatograph
the same as those published previously [21].
with a Prodigy 5 µm ODS(3) 10.0 × 250 mm
The structures of these compounds are shown
column. The mobile phase consisted of a water
and methanol gradient containing 0.2% acetic
amino-1-ethyl-6-fluoro-4-oxo-1,4-dihydro-
acid [22] at a flow rate of 3.5 mL/min. The
quinoline-3-carboxylic acid (I), and the other
diode array detector was monitored at 280 nm,
metabolites were N-formylnorfloxacin (III), N-
and metabolite concentrations were estimated
acetylnorfloxacin (IV), and desethylene-N-ace-
tylnorfloxacin (II). Two norfloxacin metabo-
lites were identified in cultures of P. guepini
trometry analyses were performed on a Hew-
during growth on corncobs dosed with nor-
WILLIAMS ET AL.: DRUG DEGRADATION IN LITTER
FIGURE 1. HPLC chromatograms (280 nm) of extracts from cultures of Pestalotiopsis guepini grown for 20 d onpoultry litter materials dosed with norfloxacin, showing norfloxacin and fungal metabolites I to IV (A: rice hulls; B:corncobs; C: pine shavings). Peaks representing compounds that were shown by mass spectrometry to be unrelatedto norfloxacin are not numbered.
floxacin (Figure 1B); the major metabolite was
The relative amounts of the norfloxacin me-
N-formylnorfloxacin (III), and the minor one
tabolites produced on poultry litter materials
was N-acetylnorfloxacin (IV). Little growth of
at 20 d were estimated from the total areas of
the fungus was observed on pine shavings, and
all the ultraviolet peaks in the HPLC chromato-
no norfloxacin metabolites were detected (Fig-
grams (Table 1). In rice-hull cultures, 18.4%
of the total consisted of 7-amino-1-ethyl-6-
FIGURE 2. Structures of norfloxacin and the 4 metabolites produced from it by Pestalotiopsis guepini during growthon rice hulls dosed with norfloxacin [21].
fluoro-4-oxo-1,4-dihydroquinoline-3-carbox-
is the predominant metabolite in broth cultures.
ylic acid (I), 0.5% was desethylene-N-acetyl-
In rice-hull cultures, more of metabolite I (7-
norfloxacin (II), 4.2% was N-formylnorfloxa-
amino-1-ethyl-6-fluoro-4-oxo-1,4-dihydro-
cin (III), and 2.3% was N-acetylnorfloxacin
quinoline-3-carboxylic acid), which has lost
(IV). In corncob cultures, 10.7% of the total
the piperazine ring, and less of metabolites II
was N-formylnorfloxacin (III), and 3.4% was
and IV were formed. In corncob cultures, me-
tabolites III (N-formylnorfloxacin) and IV (N-
acetylnorfloxacin) were found, but there were
tected in poultry litter materials are also pro-
no products requiring cleavage of the pipera-
duced in sucrose-peptone broth cultures with
zine ring. Little growth of P. guepini and no
norfloxacin [21], although N-acetylnorfloxacin
metabolites were observed on pine shavings,
WILLIAMS ET AL.: DRUG DEGRADATION IN LITTER
TABLE 1. Amounts of each of the norfloxacin metabolites produced by Pestalotiopsis guepinii during growth for20 d on poultry litter materials dosed with norfloxacin
Percentage of totalA represented by each metabolite
AMeans and standard errors from triplicate cultures, based on the total integrated areas at 280 nm of all of the identifiedmetabolite peaks plus residual norfloxacin. BND = not detected.
which contain stilbenes, resin acids, and other
quinolone, an oxonorfloxacin, a methyl ester,
compounds known to inhibit many fungi [23].
and an N-acetylethylenediamine-substituted
Another fungus, Trichoderma viride, me-
quinolone [26]. Since the known metabolites
tabolizes norfloxacin to the conjugate 4-
of fluoroquinolones are generally less active as
hydroxy-3-oxo-4-vinylcyclopent-1-enylnor-
antibacterial agents than the parent drugs [27],
floxacin when grown in sucrose-peptone broth
the fungal transformation of fluoroquinolones
[24] and to the same conjugate plus N-acetyl-
in poultry litter materials may reduce the selec-
norfloxacin when grown on rice hulls [25]. The
tive pressure on bacteria toward increased drug
mammalian metabolites of norfloxacin include
resistance. The type of litter materials used
N-acetyl- and N-formylnorfloxacin, an ethyl-
may affect the growth of fungi and the transfor-
enediamine-substituted quinolone, an amino-
CONCLUSIONS AND APPLICATIONS
1. Two common poultry litter materials, rice hulls and corncobs, served as nutrients for the
fungus Pestalotiopsis guepini and allowed it to transform added norfloxacin.
2. When grown on rice hulls dosed with norfloxacin, P. guepini produced 7-amino-1-ethyl-6-
fluoro-4-oxo-1,4-dihydroquinoline-3-carboxylic acid, N-formylnorfloxacin, N-acetylnor-floxacin, and desethylene-N-acetylnorfloxacin. When grown on corncobs, the fungus producedonly N-formylnorfloxacin and N-acetylnorfloxacin.
3. Pestalotiopsis guepini did not grow well on pine shavings, nor did it metabolize norfloxacin
REFERENCES AND NOTES
1. Endtz, H. P., G. J. Ruijs, B. van Klingeren, W. H. Jansen,
4. White, D. G., L. J. V. Piddock, J. J. Maurer, S. Zhao, V.
T. van der Reyden, and R. P. Mouton. 1991. Quinolone resistance
Ricci, and S. G. Thayer. 2000. Characterization of fluoroquinolone
in Campylobacter isolated from man and poultry following the
resistance among veterinary isolates of avian Escherichia coli.
introduction of fluoroquinolones in veterinary medicine. J. Antimi-
Antimicrob. Agents Chemother. 44:2897–2899.
5. Hofacre, C. L., A. R. de Cotret, J. J. Maurer, A. Garritty,
2. Bazile-Pham-Khac, S., Q. C. Truong, J.-P. Lafont, L. Gut-
and S. G. Thayer. 2000. Presence of fluoroquinolone-resistant coli-
mann, X. Y. Zhou, M. Osman, and N. J. Moreau. 1996. Resistanceto fluoroquinolones in Escherichia coli isolated from poultry. Anti-
forms in poultry litter. Avian Dis. 44:963–967.
microb. Agents Chemother. 40:1504–1507.
6. Cunha, B. A. 1994. The fluoroquinolones for urinary tract
3. Blanco, J. E., M. Blanco, A. Mora, and J. Blanco. 1997.
infections: A review. Adv. Ther. 11:277–296.
Prevalence of bacterial resistance to quinolones and other antimi-crobials among avian Escherichia coli strains isolated from septice-
7. Graninger, W., K. Zedtwitz-Liebenstein, H. Laferl, and H.
mic and healthy chickens in Spain. J. Clin. Microbiol. 35:2184–
Burgmann. 1996. Quinolones in gastrointestinal infections. Che-
8. Smith, A., P. M. Pennefather, S. B. Kaye, and C. A. Hart.
19. Bacon, C. W., and D. Burdick. 1977. Growth of fungi in
2001. Fluoroquinolones: place in ocular therapy. Drugs 61:747–
broiler houses. Poult. Sci. 56:653–661.
20. Sˇkrinjar, M., M. Ristic´, and Z. Grbic´. 1995. Contamination
9. Laczay, P., G. Semje´n, G. Nagy, and J. Lehel. 1998. Com-
of broiler chicken’s mash and litter with moulds, aflatoxins, ochra-
parative studies on the pharmacokinetics of norfloxacin in chickens,
toxin A and zearalenone. Acta Vet. Hung. 43:117–124.
turkeys and geese after a single oral administration. J. Vet. Pharma-
21. Parshikov, I. A., T. M. Heinze, J. D. Moody, J. P. Freeman,
A. J. Williams, and J. B. Sutherland. 2001. The fungus Pestaloti-
10. Sumano, L. H., C. L. Ocampo, G. W. Brumbaugh, and R. opsis guepini as a model for biotransformation of ciprofloxacin
E. Lizarraga. 1998. Effectiveness of two fluoroquinolones for the
and norfloxacin. Appl. Microbiol. Biotechnol. 56:474–477.
treatment of chronic respiratory disease outbreak in broilers. Br.
22. Parshikov, I. A., J. P. Freeman, J. O. Lay, R. D. Beger, A.
J. Williams, and J. B. Sutherland. 1999. Regioselective transforma-
11. Al-Mustafa, Z. H., and M. S. Al-Ghamdi. 2000. Use of
tion of ciprofloxacin to N-acetylciprofloxacin by the fungus Mucor
norfloxacin in poultry production in the eastern province of Saudi
ramannianus. FEMS Microbiol. Lett. 177:131–135.
Arabia and its possible impact on public health. Int. J. Environ. Health Res. 10:291–299.
23. Celimene, C. C., J. A. Micales, L. Ferge, and R. A. Young.
1999. Efficacy of pinosylvins against white-rot and brown-rot
12. Kelley, T. R., O. C. Pancorbo, W. C. Merka, and H. M.
Barnhart. 1998. Antibiotic resistance of bacterial litter isolates. Poult. Sci. 77:243–247.
24. Parshikov, I. A., J. D. Moody, J. P. Freeman, J. O. Lay,
A. J. Williams, T. M. Heinze, and J. B. Sutherland. 2002. Formation
13. Joseph, S. W., J. R. Hayes, L. L. English, L. E. Carr,
of conjugates from ciprofloxacin and norfloxacin in cultures of
and D. D. Wagner. 2001. Implications of multiple antimicrobial-
Trichoderma viride. Mycologia 94:1–5.
resistant enterococci associated with the poultry environment. FoodAddit. Contam. 18:1118–1123.
25. Williams, A. J., I. A. Parshikov, J. D. Moody, T. M. Heinze,
J. P. Freeman, and J. B. Sutherland. 2001. The metabolism of two
14. Eriksson de Rezende, C. L., E. T. Mallinson, N. L. Tablante,
antibacterial agents, norfloxacin and sarafloxacin, by the saprobic
R. Morales, A. Park, L. E. Carr, and S. W. Joseph. 2001. Effect
fungus Trichoderma viride during growth on rice hulls. Page 622
of dry litter and airflow in reducing Salmonella and Escherichia
in Abstr. Am. Soc. Microbiol. 101st Gen. Mtg., Orlando, FL. coli populations in the broiler production environment. J. Appl. Poult. Res. 10:245–251.
26. Pauliukonis, L. T., D. G. Musson, and W. F. Bayne. 1984.
Quantitation of norfloxacin, a new antibacterial agent in human
15. Payne, J. B., E. C. Kroger, and S. E. Watkins. 2002. Evalua-
plasma and urine by ion-pair reverse-phase chromatography. J.
tion of litter treatments on Salmonella recovery from poultry litter.
27. Zeiler, H.-J., U. Petersen, W. Gau, and H. J. Ploschke.
16. Lu, J., S. Sanchez, C. Hofacre, J. J. Maurer, B. G. Harmon,
1987. Antibacterial activity of the metabolites of ciprofloxacin
and M. D. Lee. 2003. Evaluation of broiler litter with reference to
and its significance in the bioassay. Arzneim.-Forsch. Drug Res.
the microbial composition as assessed by using 16S rRNA and
functional gene markers. Appl. Environ. Microbiol. 69:901–908.
17. Lovett, J., J. W. Messer, and R. B. Read. 1971. The mi-
croflora of southern Ohio poultry litter. Poult. Sci. 50:746–751. Acknowledgments
18. Lovett, J. 1972. Toxigenic fungi from poultry feed and
We thank C. E. Cerniglia, J. P. Freeman, and F. Rafii for helpful
discussions and comments on the manuscript.
Great-West’s Drug Prior Authorization Great-West Life’s prior authorization process is designed to provide an effective approach to managing claims for specific prescription drugs. Approval for coverage of certain prescription drugs is required in order to provide plan members with coverage for appropriate drug treatment, and keep your drug plan affordable. Below is a current listing of dr
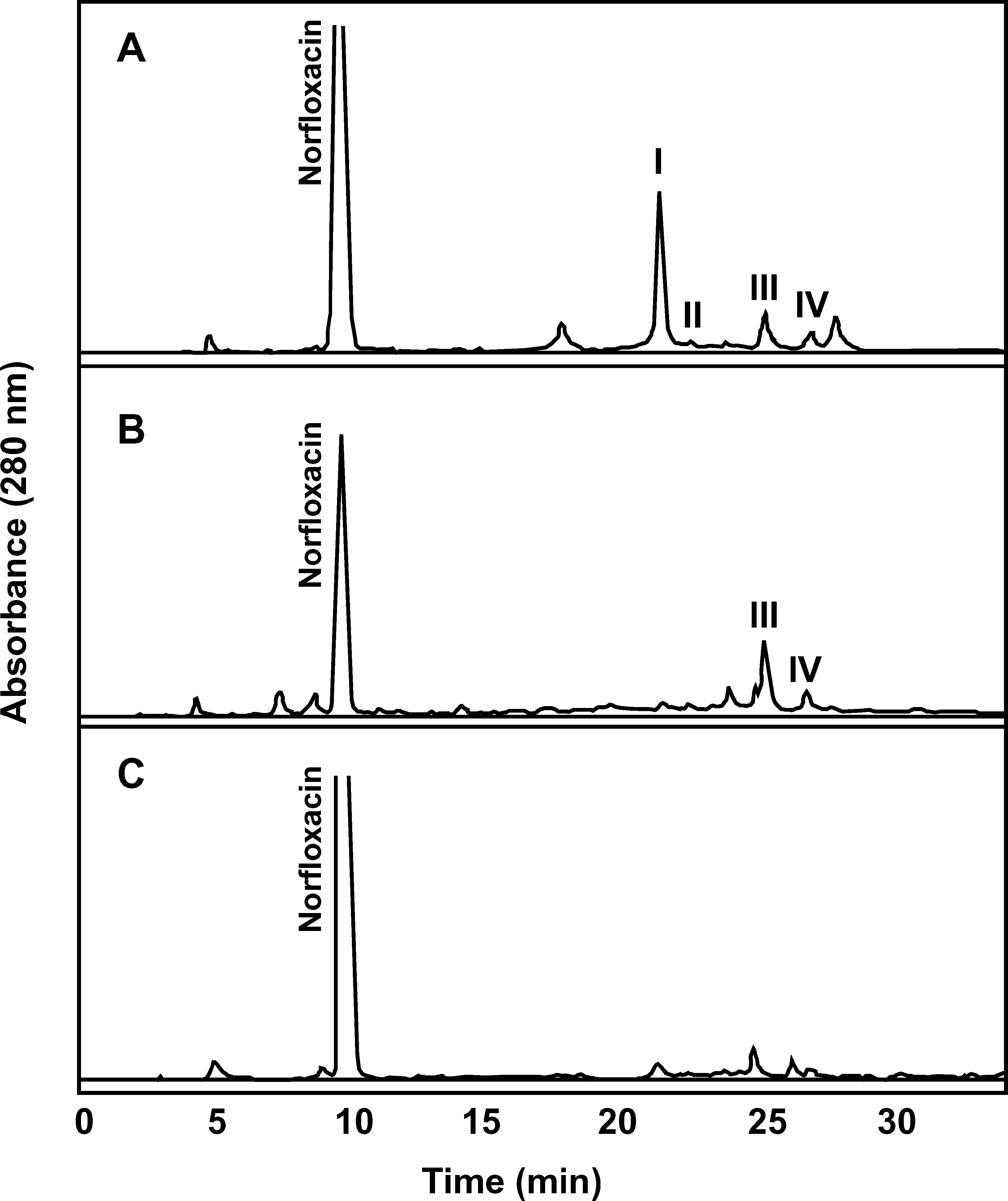 WILLIAMS ET AL.: DRUG DEGRADATION IN LITTER
FIGURE 1. HPLC chromatograms (280 nm) of extracts from cultures of Pestalotiopsis guepini grown for 20 d onpoultry litter materials dosed with norfloxacin, showing norfloxacin and fungal metabolites I to IV (A: rice hulls; B:corncobs; C: pine shavings). Peaks representing compounds that were shown by mass spectrometry to be unrelatedto norfloxacin are not numbered.
WILLIAMS ET AL.: DRUG DEGRADATION IN LITTER
FIGURE 1. HPLC chromatograms (280 nm) of extracts from cultures of Pestalotiopsis guepini grown for 20 d onpoultry litter materials dosed with norfloxacin, showing norfloxacin and fungal metabolites I to IV (A: rice hulls; B:corncobs; C: pine shavings). Peaks representing compounds that were shown by mass spectrometry to be unrelatedto norfloxacin are not numbered.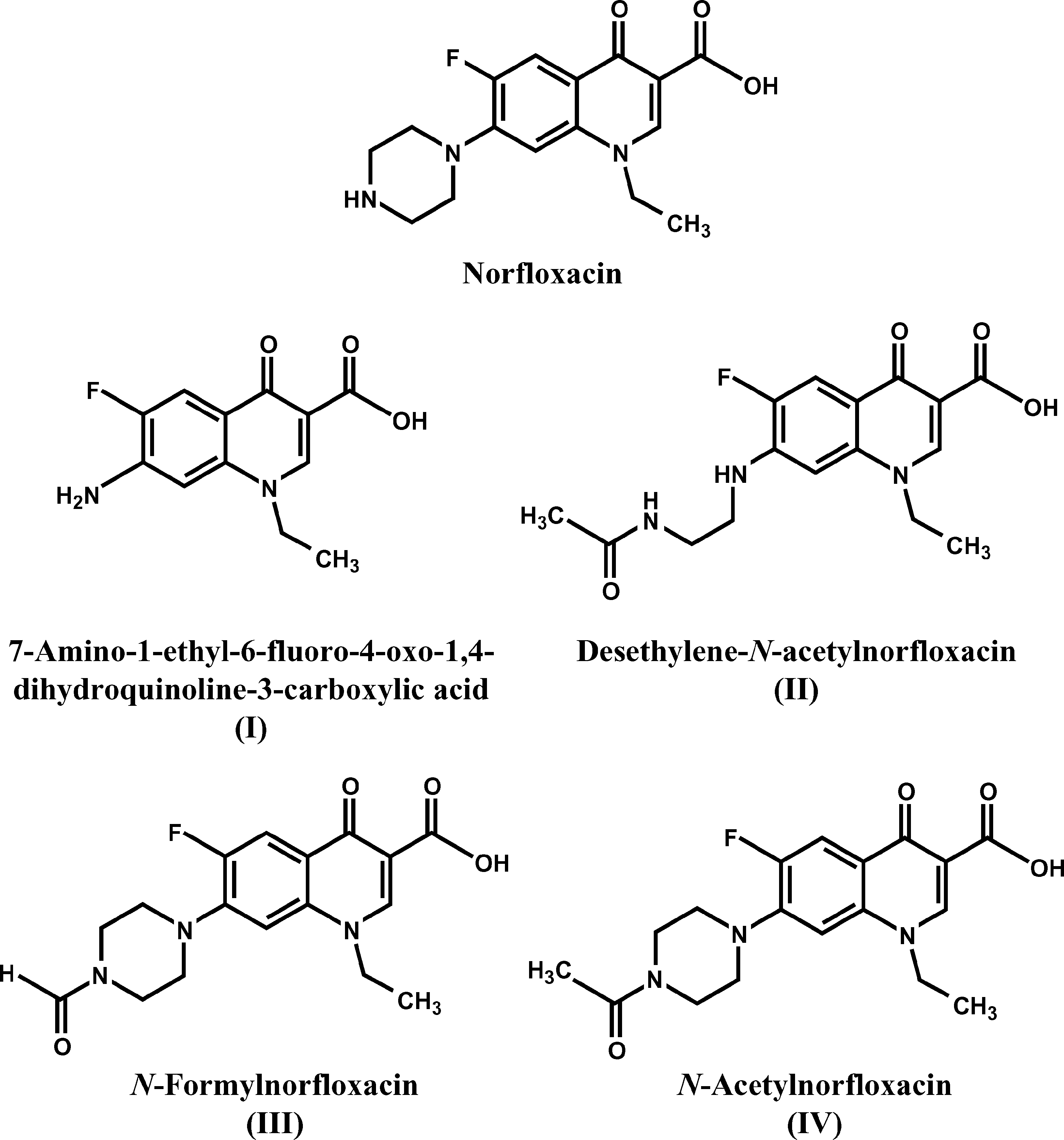 FIGURE 2. Structures of norfloxacin and the 4 metabolites produced from it by Pestalotiopsis guepini during growthon rice hulls dosed with norfloxacin [21].
FIGURE 2. Structures of norfloxacin and the 4 metabolites produced from it by Pestalotiopsis guepini during growthon rice hulls dosed with norfloxacin [21].