Do you want to buy antibiotics online without prescription? https://buyantibiotics24h.net/ - This is pharmacy online for you!
Doi:10.1016/j.febslet.2006.05.036
TC1(C8orf4) is upregulated by IL-1b/TNF-a and enhances
proliferation of human follicular dendritic cells
Youngmi Kima,1, Jungtae Kima,1, Juhee Parka, Seunghyun Banga, Yusun Junga,
Jongseon Choeb, Kyuyoung Songc, Inchul Leed,*
a Asan Institute for Life Sciences, University of Ulsan, College of Medicine, Seoul 138-736, Republic of Korea
b Department of Microbiology and Immunology, Kangwon National University, College of Medicine, Chunchon, Kangwon-Do 200-701,
c Department of Biochemistry and Molecular Biology, University of Ulsan, College of Medicine, Seoul 138-736, Republic of Korea
d Department of Pathology, University of Ulsan, College of Medicine, 388-1 Poongnap-Dong, Songpa-Gu, Seoul 138-736, Republic of Korea
Received 29 March 2006; revised 9 May 2006; accepted 15 May 2006
that has been widely implicated in regulating cell proliferation
Follicular dendritic cells (FDC) play crucial roles in
in cancers and development . TC1 is a small protein in
immune regulation. TNF-a has been shown to be essential tothe FDC network. However, the molecular regulation of FDC
vertebrates which enhances the signaling pathway by
proliferation has not been characterized. Here, we show that
relieving the antagonistic function of Chibby on b-catenin
TC1(C8orf4), a novel positive regulator of the Wnt/b-catenin
mediated transcription TC1 enhances cancer cell prolifer-
pathway in vertebrates, is upregulated by IL-1b and TNF-a in
ation and invasiveness The expression of TC1 cor-
the human FDC-like line HK. TC1 enhances HK cell prolifera-
related with aggressive biological behavior of gastric cancer
tion, while TC1-knockdown inhibits the proliferation induced by
and with poor survival . However, the transcriptional reg-
IL-1b, suggesting a role of TC1 as a regulator of FDC prolifer-
ulation of TC1 has remained to be investigated.
ation. The regulation by pro-inflammatory cytokines suggests
In this study, we investigated the biological effects of TNF-a
that TC1 might be implicated in linking local inflammation to
and IL-1b on TC1 expression. Human FDC-like line HK
immune response by stimulating FDC. Ó
was used for the investigation because of the crucial roles of
2006 Federation of European Biochemical Societies. Published
by Elsevier B.V. All rights reserved.
FDCs in the immune system and the requirement of TNF-afor the proliferation. We were also interested in the potential
Keywords: TC1(C8orf4); NF-kB; IL-1b; TNF-a;
biological role of TC1 in non-neoplastic primary culture cells.
Here, we show that IL-1b and TNF-a upregulate TC1
expression in a NF-jB-dependent manner. TC1 upregulatedb-catenin target genes in HK cells as in cancer cells. TC1 en-hanced the proliferation of HK cells while TC1-knockdowninhibited the proliferation induced by IL-1b, suggesting a bio-
logical role of TC1 as a major regulator of FDC proliferation. The regulation of TC1 by pro-inflammatory cytokines suggests
Follicular dendritic cells (FDC) play crucial roles in germinal
that it might be implicated in linking local inflammation to B
center formation and B cell differentiation (reviewed in
FDCs display elaborate cytoplasmic processes which pres-ent native antigens on the surface in the form of immune-complexes through Fc receptors and complement receptors.
FDCs organize mature lymphoid follicles collaborating withB and T cells . TNF-a has been shown to be essential
FDC-like cell line HK was established from human tonsils and
to the development of the FDC network in germinal centers
grown in RPMI 1640 supplemented with 10% fetal bovine serum,
. Pro-inflammatory cytokines TNF-a and IL-1b activate
2 mM L-glutamine, and 80 lg/ml gentamicin in a humidified atmo-
IKKs, which target IjB for polyubiquitination and subsequent
sphere of 5% CO2 at 37 °C . Experiments were done using
degradation by proteasome, thereby releasing NF-jB. The acti-
HK cells of 9–13 passages. Human recombinant IL-1b and TNF-a
vation of NF-jB has been widely implicated in regulation of
were purchased commercially (Sigma, St. Louis, MO), and appliedat 20 U/ml media and 10 ng/ml, respectively. Sulindac sulfide and
immunity, inflammation, and apoptosis . However, it has
PDTC (pyrrolidine dithiocarbamate) were also purchased (Sigma),
remained to be elucidated how NF-jB activation enhances
Recently, we have reported that TC1(C8orf4) functions as a
positive regulator of the Wnt/b-catenin signaling pathway
HA-TC1 construct in pcDNA3 vector (Invitrogen, Carldbad, CA)
was described previously Lenti-TC1, a full length TC1-expressinglentiviral vector was constructed by inserting TC1 ORF into lentiM1.2
*Corresponding author. Fax: +82 2 472 7898.
vector (VectorCore A Inc., Deajeon, Korea), which had mCMV pro-
moter for the expression and eGFP-Zeocin fusion protein under IRESpromoter. LentiM1.4-eGFP vector was used as control. Lenti-siTC1,
1 Both authors contributed equally to this work.
a shRNA-expression lentiviral vector for targeting TC1 gene, was also
0014-5793/$32.00 Ó 2006 Federation of European Biochemical Societies. Published by Elsevier B.V. All rights reserved. doi:10.1016/j.febslet.2006.05.036
Y. Kim et al. / FEBS Letters 580 (2006) 3519–3524
constructed by inserting synthetic oligonucleotide, 50-CGGAATT-
CACACAGACCAAGAATCACTAGAAAGttcaagagaCTTTCTAGTG-ATTCTTGGTCTGTGTTTTTTGATATCTAGACA-30
letters represent sense and antisense sequences of TC1, small lettersrepresent an insert for loop, and the 5 Ts represent a transcription stop
signal) into the EcoRI–XbaI site of shLenti2.4G vector (VectorCore A
Inc.). The siRNA sequence was previously shown to downregulateTC1 expression efficiently and specifically . The control con-
sisted of scrambled shRNA sequences (50-AATCGCATAGCG-TATGCCGTT-30) inserted into the same vector. Lentiviruses were
prepared according to the standard protocol
2.3. Real-time and semiquantitative RT-PCR
TC1 expression levels were measured using real-time PCR analysis.
HK cells were treated with IL-1b, TNF-a, sulindac and/or PDTC as
indicated. Total RNA was extracted using Trizol reagent (Invitrogen)
and cDNA was synthesized using Superscript II reverse transcriptase
(Invitrogen). Quantitative PCR was performed using a continuousfluorescence detecting thermal cycler ABI PRISMÒ 7000 Sequence
Detection System (ABI, Foster city, CA), and a SYBRÒ Green real-
time PCR master mix (Toyobo, Osaka, Japan). Measurements weredone in triplicate using b-actin gene as an endogenous control. For
the statistical analysis, analysis of variance (ANOVA) test was applied
using the SPSS software (SPSS Inc. Chicago, IL).
The expressions of TC1 and/or b-catenin target genes were analyzed
after Lenti-TC1 or Lenti-siTC1 transfection using corresponding con-
trol vectors and b-actin as loading control. Semiquantitative RT-PCR
was done as described previously using primers for TC1, 50-
caagccatcatcatgtccac-30 and 50-gttgcccacggctttcttac-30; c-Myc, 50-tgctc-
catgaggagacacc-30 and 50-ctctgaccttttgccaggag-30; c-MET, 50-gcatttt-tacggacccaatc-30
and 50-gctgcaaagctgtggtaaac-30; Cyclin D1, 50-
tcctctccaaaatgccagag-30 and 50-ggcggattggaaatgaact-30; and b-actin
50-ggcacccagcacaatgaag-30 and 50-gccgatccacacggagtact-30.
Fig. 1. (A) TC1 upregulation by IL-1b in HK cells. After the treatment
of 20 U/ml IL-1b, TC1 mRNA levels were measured by SYBR-green
HK cells grown on cover slips were immunostained using rabbit
real-time PCR in quadruplicate using b-actin as internal control. The
anti-Rel A antiserum (Santa Cruz Biotechnology, Santa Cruz, CA).
TC1 expression is enhanced in an hour, reaching more than 10 times of
After washing with PBS, FITC-labeled anti-rabbit second antibody
the basal level in 3 h. (B) TC1 upregulation by IL-1b and TNF-a. TC1
(Jackson Immunoresearch, West Grove, PN) was applied. DNA stain-
levels were measured at 6 h after the treatment of 20 U/ml IL-1b and/
ing was done with DAPI (Sigma). Cells were viewed using an Olympus
or 10 ng/ml TNF-a. TNF-a also enhances TC1 but to a lesser degree
BX51 fluorescence microscope. For control, the primary antibody was
than IL-1b does. IL-1b and TNF-a together show a synergistic effect.
and NF-jB inhibitor PDTC Both sulindac sulfide, an ac-
The proliferation of HK cells with or without treatment of IL-1b
tive metabolite of sulindac, and PDTC were applied at rela-
and/or TNF-a was analyzed using WST-1 proliferation assay kit
tively low levels, 50 lM, which did not affect cell growth or
according to the manufacturer’s instruction (Roche, Mannheim, Ger-
damage (data not shown). After 3 h, TC1 levels were measured
many). Lenti-TC1 or Lenti-siTC1-transfected HK cells were analyzed
and compared with untreated control. Both sulindac and
using corresponding controls. Measurements were done in quadrupli-cate at 24, 48, and 72 h after cytokine treatments, and fold changes in
PDTC inhibited IL-1b-induced TC1 upregulation significantly
comparison with controls were analyzed using ANOVA test.
(P < 0.001), suggesting that the upregulation was NF-jB-dependent ). Given the prompt upregulation of TC1after the cytokine treatments, our data suggested TC1 as one
3.1. IL-1b and TNF-a enhances TC1 expression in HK cells
3.3. NF-kB activation by IL-1b in HK cells
TC1 expression was at low level in HK cells normally. Upon
It has been shown that TNF-a activates NF-jB in HK cells
IL-1b treatment, TC1 expression was upregulated promptly,
. However, the effect of IL-1b on the NF-jB activation has
reaching more than 10 times of the basal level in 3 h as mea-
not been reported in HK cells. Upon the activation by the cyto-
sured by real-time PCR A). After 24 h, TC1 expression
kines, NF-jB is released from IjB to be translocated into the
decreased to remain at a considerably high level compared to
nucleus . Then, NF-jB regulates target genes including
the untreated control. Under the experimental conditions,
IjB and, thereby, the transcriptional activity of NF-jB is re-
IL-1b upregulated TC1 much more than TNF-a did
duced again rather rapidly upon the negative feedback by en-
IL-1b and TNF-a enhanced the proliferation syner-
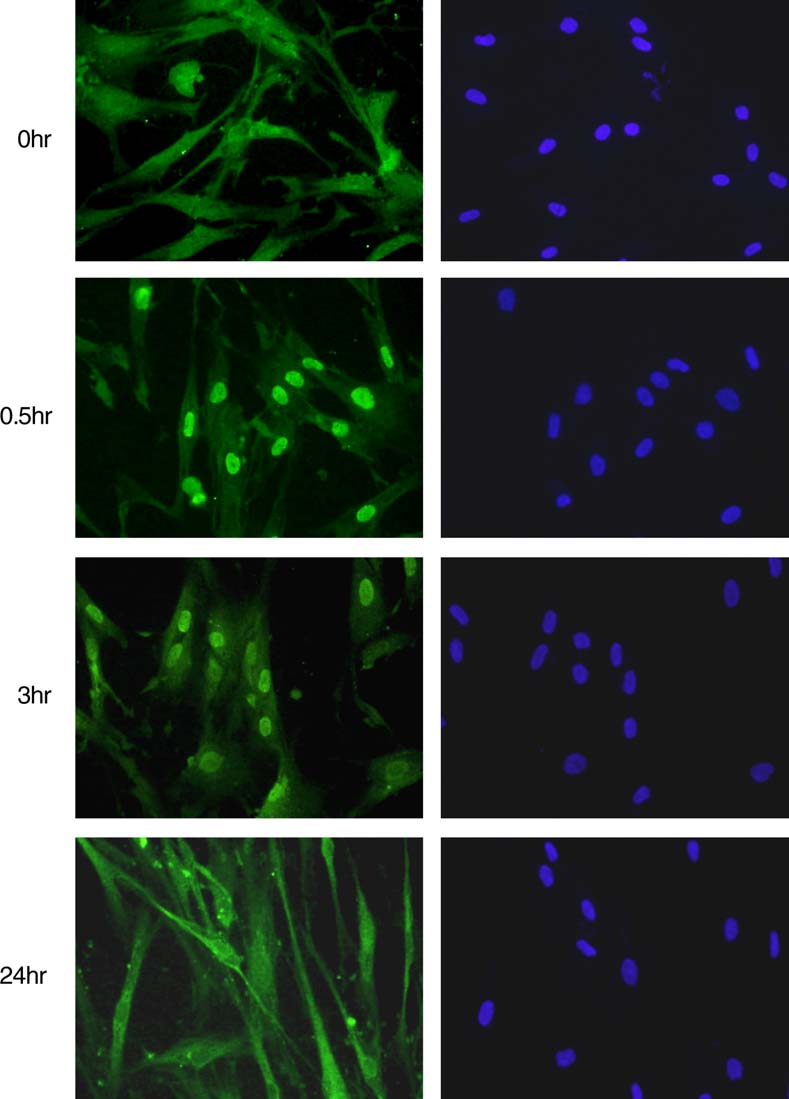
immunofluorescence staining for Rel A, a subunit of NF-jB. Upon IL-1b treatment, the nuclear RelA immunostaining
3.2. IL-1b-induced TC1 upregulation is NF-kB-dependent
was detected in 30 min, and then decreased mildly after 3 h
The upregulation of TC1 by IL-1b and TNF-a suggested
(). The nuclear immunostaining largely disappeared in
that TC1 might be a target gene of NF-jB. To test the hypoth-
24 h. Our data showed that IL-1b also activated NF-jB in
esis, we then analyzed the effects of IKK inhibitor sulindac
Y. Kim et al. / FEBS Letters 580 (2006) 3519–3524
3.4. TC1 upregulates b-catenin target genes in HK cells
Previously, we have shown that TC1 upregulates b-catenin
target genes in cancer cells Because TC1 expression level
was quite low in HK line, a gain-of-function study by introduc-ing exogenous TC1 would have been appropriate. However, HK
cells are resistant to conventional gene transfer methods (data
not shown). Thus, we constructed the full length TC1 in lentiviral
vector to facilitate the gene transfer. HK cells were transfectedwith lentiviral constructs effectively so that most cells expressed
GFP in 24 h (data not shown). TC1-transfected HK cells showed
markedly upregulated TC1 expression in comparison with con-
trol vector-transfected cells as measured by real-time PCR
Fig. 2. Effect of sulindac and PDTC on IL-1b-induced TC1 upregu-
The expressions of cyclin D1, c-Myc, and c-Met were
lation. HK cells were treated with 20 U/ml IL-1b, 50 lM sulindac
enhanced significantly compared to the control as
sulfide, and/or 50 lM PDTC. After 3 h, TC1 levels were measured by
measured by semiquantitative RT-PCR . They are b-catenin
SYBR-green real-time PCR in quadruplicate using b-actin as internal
target genes and are implicated in cell proliferation (reviewed in
control. Sulindac and PDTC inhibited TC1 upregulation by IL-1bin a dose-dependent manner, suggesting an NF-jB-dependent
). Our data suggest that TC1 functions as a positive reg-
ulator of the Wnt/b-catenin signaling pathway in HK cells also.
Fig. 3. IL-1b induces nuclear translocation of NF-jB activation by in HK cells. HK cells grown on cover slips were immunostained using rabbit anti-Rel A antiserum. After washing with PBS, FITC-labeled anti-rabbit second antibody was applied. DNA staining was done using DAPI, and cellswere viewed using Olympus BX51 fluorescence microscope.
Y. Kim et al. / FEBS Letters 580 (2006) 3519–3524
Fig. 4. TC1 upregulates b-catenin target genes in HK cells. (A) TC1 expression in Lenti-TC1 or control vector-transfected cells. The expression wasmeasured by real-time PCR in quadruplicate using b-actin as internal control. (B) Semiquantitative RT-PCR for b-catenin target genes cyclin D1, c-Myc and c-Met in Lenti-TC1-transfected HK cells and control vector-transfected cells. Densitometric values were normalized using b-actin asloading control.
We then investigated the effect of TC1 transfection with or
The upregulation of cyclin D1, c-Myc, and c-Met suggested
without IL-1b treatment. TC1-transfected cells showed signif-
that TC1 might enhance the proliferation of HK cells. We first
icant enhancement of proliferation in 72 h compared to the
analyzed the effects of IL-1b and TNF-a on HK cell prolifer-
control vector-transfected cells (P < 0.001, The
ation. TNF-a was shown to enhance the proliferation of HK
enhancement by TC1 was higher than that by IL-1b alone in
cells ; however, the role of IL-1b has not been reported.
72 h (P < 0.05). TC1 and IL-1b showed an additive effect on
IL-1b and TNF-a showed similar enhancing effects on the pro-
liferation compared to untreated HK cells (P < 0.001,
To investigate the normal requirements of TC1 in HK cell
A). IL-1b and TNF-a together showed a significant syn-
proliferation, we next analyzed the effect of TC1 knockdown
ergistic effect (P < 0.001), enhancing the proliferation continu-
on HK cell proliferation. Lenti-siRNA was constructed using
a siRNA sequence that was shown to downregulate TC1
Absorbance (A Absorbance (A Time (hr) Time (hr) Expression fold 0.25 Absorbance (A 1.5 Time (hr)
Fig. 5. TC1 and cytokine effects on HK cell proliferation. (A) Effects of IL-1b and/or TNF-a on HK cell proliferation. 20 U/ml IL-1b and/or 10 ng/ml TNF-a was applied to HK cells, and the proliferation was analyzed using WST-1 proliferation assay kit (Roche). Measurements were done inquadruplicate at 24, 48, and 72 h after cytokine treatments. Fold changes were analyzed in comparison with controls using ANOVA test. (B) Theproliferation of TC1-Lenti- and control vector-transfected HK cells with or without IL-1b was measured similarly. (C) TC1 knockdown using Lenti-siTC1 transfection compared to the control vector-transfected HK cells. TC1 was measured by real-time PCR in quadruplicate using b-actin asinternal control. (D) The proliferation of Lenti-siTC1- and control vector-transfected HK cells with or without IL-1b was measured similarly.
Y. Kim et al. / FEBS Letters 580 (2006) 3519–3524
expression efficiently and specifically Lenti-siTC1
downregulated endogenous TC1 significantly compared tothe control vector (C). TC1-knockdown inhibited the
[1] Kosco-Vilbois, M.H. (2003) Are follicular dendritic cells really
proliferation compared to the control vector-transfected cells
good for nothing? Nat. Rev. Immunol. 3, 764–769.
[2] Park, C.S. and Choi, Y.S. (2005) How do follicular dendritic cells
(P < 0.001, Lenti-siTC1 inhibited the IL-1b-induced
interact intimately with B cells in the germinal centre? Immunol-
enhancement significantly (P < 0.001). Together, our data sug-
gest that TC1 is a major regulator of FDC proliferation, and
[3] Klaus, G.G., Humphery, J.H., Kunkl, A. and Dongworth, D.W.
the enhancing effect of IL-1b might be TC1-dependent.
(1980) The follicular dendritic cell: its role in antigen presentationin the generation of immunological memory. Immunol. Rev. 53,3–28.
[4] Ansel, K.M., Ngo, V.N., Hyman, P.L., Luther, S.A., Forster, R.,
Sedgwick, J.D., Browning, J.L., Lipp, M. and Cyster, J.G. (2000)A chemokine-driven positive feedback loop organizes lymphoidfollicles. Nature 406, 309–314.
The regulation of FDC proliferation is crucial for B-cell
[5] Pasparakis, M., Alexopoulou, L., Episkopou, V. and Kollias, G.
mediated immune response. TNF-a is essential for the devel-
(1996) Immune and inflammatory responses in TNF alpha-
opment of FDC network in the germinal center. However, it
deficient mice: a critical requirement for TNF alpha in the
has remained to be elucidated how NF-jB activation by
formation of primary B cell follicles, follicular dendritic cell
TNF-a enhances the proliferation of FDCs. Our data suggest
networks and germinal centers, and in the maturation of thehumoral immune response. J. Exp. Med. 184, 1397–1411.
that TC1 might be a NF-jB-dependent regulator of FDC pro-
[6] Park, S.M., Park, H.Y. and Lee, T.H. (2003) Functional effects of
liferation. The suggested role of TC1 is compatible with the its
TNF-alpha on a human follicular dendritic cell line: persistent
proposed function as a positive regulator of the Wnt/b-catenin
NF-kappa B activation and sensitization for Fas-mediated
apoptosis. J. Immunol. 171, 3955–3962.
[7] Chen, Z.J. (2005) Ubiquitin signalling in the NF-kappaB path-
TC1 was upregulated promptly by IL-1b/TNF-a and was
inhibited by IKK/NF-jB inhibitors, suggesting TC1 as one
[8] Schmitz, M.L., Mattioli, I., Buss, H. and Kracht, M. (2004) NF-
of target genes of NF-jB. Both IL-1b and TNF-a activated
kappaB: a multifaceted transcription factor regulated at several
NF-jB and enhanced the proliferation of HK cells synergisti-
cally. However, TC1 upregulation by IL-1b was significantly
[9] Karin, M., Yamamoto, Y. and Wang, Q.M. (2004) The IKK NF-
kappa B system: a treasure trove for drug development. Nat. Rev.
higher than by TNF-a. Together, our data suggest a significant
[10] Jung, Y., Bang, S., Choi, K., Kim, E., Kim, J., Park, J., Koo, H.,
Both gain- and loss-of-function analyses suggested that TC1
Moon, R.T., Song, K. and Lee, I. (2006) TC1 (C8orf4) enhances
was implicated in the regulation of HK cell proliferation. TC1
the Wnt/beta-catenin pathway by relieving antagonistic activity ofChibby. Cancer Res. 66, 723–728.
is present in vertebrates, and is tightly regulated at both tran-
[11] Giles, R.H., van Es, J.H. and Clevers, H. (2003) Caught up in a
scriptional and post-translational levels Nonetheless,
Wnt storm: Wnt signaling in cancer. Biochim. Biophys. Acta
TC1-knockdown downregulated HK cell proliferation and
inhibited IL-1b-induced proliferation significantly, suggesting
[12] Nelson, W.J. and Nusse, R. (2004) Convergence of Wnt, beta-
that TC1 is a major regulator of FDC proliferation. The regu-
catenin, and cadherin pathways. Science 303, 1483–1487.
[13] Moon, R.T., Kohn, A.D., De Ferrari, G.V. and Kaykas, A.
lation of TC1 by pro-inflammatory cytokines such as IL-1b
(2004) WNT and beta-catenin signalling: diseases and therapies.
and TNF-a suggests that TC1 might be implicated in linking
local inflammation to immune response by stimulating FDCs.
[14] Chua, E.L., Young, L., Wu, W.M., Turtle, J.R. and Dong, Q.
HK cells express FDC markers such as HJ2, GP93, 3C8,
(2000) Cloning of TC-1 (C8orf4), a novel gene found to beoverexpressed in thyroid cancer. Genomics 69, 342–347.
8D6, DRC1, and KIM4 (reviewed in ). Functionally, HK
[15] Nicod, M., Michlig, S., Flahaut, M., Salinas, M., Fowler Jaeger,
cells preferentially bind germinal center B cells, providing them
N., Horisberger, J.D., Rossier, B.C. and Firsov, D. (2002) A novel
with survival signals, while the majority of unbound B cells un-
vasopressin-induced transcript promotes MAP kinase activation
dergo apoptosis promptly. However, functional analyses of
and ENaC downregulation. EMBO J. 21, 5109–5117.
individual genes in FDC-like cells have been restricted, because
[16] Takemaru, K., Yamaguchi, S., Lee, Y.S., Zhang, Y., Carthew,
R.W. and Moon, R.T. (2003) Chibby, a nuclear beta-catenin-
they are refractory to gene modulations using conventional
associated antagonist of the Wnt/Wingless pathway. Nature 422,
gene transfer methods. Our data showed that lentiviral vectors
could be applied to the functional analysis of genes of interest
[17] Sunde, M., McGrath, K.C., Young, L., Matthews, J.M., Chua,
E.L., Mackay, J.P. and Death, A.K. (2004) TC-1 is a noveltumorigenic and natively disordered protein associated with
Our data suggest that TC1 might link NF-jB and Wnt/b-
thyroid cancer. Cancer Res. 64, 2766–2773.
catenin pathways. It has been shown that b-catenin inhibits
[18] Kim, B., Koo, H., Yang, S., Bang, S., Jung, Y., Kim, J., Kim, Y.,
NF-jB activity in cancers and intestinal epithelial cells
Park, J., Moon, R.T., Song, K., and Lee, I. TC1(C8orf4)
However, no positive link between the pathways has
correlates with Wnt/b-catenin target genes and aggressive biolog-
been reported so far. The crosstalk might have a significant
ical behavior of gastric cancer. Clin Cancer Res. (in press).
[19] Kim, H.S., Zhang, X. and Choi, Y.S. (1994) Activation and
implication in cancers. Both NF-jB and Wnt/b-catenin
proliferation of follicular dendritic cell-like cells by activated T
pathways have been implicated in various cancers.
lymphocytes. J. Immunol. 153, 2951–2961.
[20] Lee, S., Choi, K., Ahn, H., Song, K., Choe, J. and Lee, I. (2005)
Acknowledgements: We appreciate Randall T. Moon for critical read-
TuJ1 (class III b-tubulin) expression suggests dynamic redistri-
ing of the manuscript. We also appreciate Chan-Sik Park for the dis-
bution of follicular dendritic cells in lymphoid tissue. Eur. J. Cell
cussion regarding IL-1b effect and gene transfer to HK cells. This
work was supported by the Korean Ministry of Science and Technol-
[21] Dull, T., Zufferey, R., Kelly, M., Mandel, R.J., Nguyen, M.,
ogy’s National Research Laboratory Project Grant (M10400000305-
Trono, D. and Naldini, L. (1998) A third-generation lentivirus
05J0000-30510) and Molecular and Cellular Function Discovery
vector with a conditional packaging system. J. Virol. 72, 8463–
Project Grant (M10401000003-05N0100-00310).
Y. Kim et al. / FEBS Letters 580 (2006) 3519–3524
[22] Kim, B., Bang, S., Lee, S., Kim, S., Choi, K., Lee, C., Lee, S.G.,
[27] Christensen, J.G., Burrows, J. and Salgia, R. (2005) c-Met as a
Lee, K., Lee, Y., Kim, Y., Kim, S., Yeom, Y., Yu, H., Song, K.
target for human cancer and characterization of inhibitors for
and Lee, I. (2003) Expression profiling and subtype-specific
therapeutic intervention. Cancer Lett. 225, 1–26.
expression of stomach cancer. Cancer Res. 63, 8248–8255.
[28] Deng, J., Miller, S.A., Wang, H.Y., Xia, W., Wen, Y., Zhou, B.P.,
[23] Yamamoto, Y., Yin, M.J., Lin, K.M. and Gaynor, R.B. (1999)
Li, Y., Lin, S.Y. and Hung, M.C. (2002) Beta-catenin interacts
Sulindac inhibits activation of the NF-jB pathway. J. Biol. Chem.
with and inhibits NF-kappa B in human colon and breast cancer.
[24] Liu, S.F., Ye, X. and Malik, A.B. (1999) Inhibition of NF-jB
[29] Sun, J., Hobert, M.E., Duan, Y., Rao, A.S., He, T.C., Chang,
activation by pyrrolidine dithiocarbamate prevents in vivo
E.B. and Madara, J.L. (2005) Crosstalk between NF-kappaB and
expression of proinflammatory genes. Circulation 100, 1330–
beta-catenin pathways in bacterial-colonized intestinal epithelial
cells. Am. J. Physiol. Gastrointest. Liver Physiol. 289, G129–
[25] Musgrove, E.A. (2006) Cyclins: roles in mitogenic signaling and
oncogenic transformation. Growth Factors 24, 13–19.
[30] Karin, M. and Greten, F.R. (2005) NF-kappaB: linking inflam-
[26] Gallant, P. (2005) Myc, cell competition, and compensatory
mation and immunity to cancer development and progression.
proliferation. Cancer Res. 65, 6485–6487.
Drugs & Supplements: Vitamin B12 (Print Version) Vitamin B12 URL of this page: http://www.nlm.nih.gov/medlineplus/druginfo/natural/patient-vitaminb12.html Background Vitamin B12 is an essential water soluble vitamin that is commonly found in a variety of foods such as fish, shellfish,meats, and dairy products. Vitamin B12 is frequentl
Science in China Ser. D Earth Sciences 2004 Vol. 47 No. 1 30—36 Palynological record during the Pleistocene (between 1.05 Ma and 0.36 Ma) from ODP1144, northern South China Sea 1. Institute of Geology and Paleontology, Chinese Academy of Sciences, Nanjing 210008, China;2. Institute of Botany, Chinese Academy of Sciences, Beijing 100093, China;3. Key Laboratory of Marine Geology, Ministry
 Y. Kim et al. / FEBS Letters 580 (2006) 3519–3524
3.4. TC1 upregulates b-catenin target genes in HK cells
Previously, we have shown that TC1 upregulates b-catenin
target genes in cancer cells Because TC1 expression level
was quite low in HK line, a gain-of-function study by introduc-ing exogenous TC1 would have been appropriate. However, HK
cells are resistant to conventional gene transfer methods (data
not shown). Thus, we constructed the full length TC1 in lentiviral
vector to facilitate the gene transfer. HK cells were transfectedwith lentiviral constructs effectively so that most cells expressed
GFP in 24 h (data not shown). TC1-transfected HK cells showed
markedly upregulated TC1 expression in comparison with con-
trol vector-transfected cells as measured by real-time PCR
Fig. 2. Effect of sulindac and PDTC on IL-1b-induced TC1 upregu-
The expressions of cyclin D1, c-Myc, and c-Met were
lation. HK cells were treated with 20 U/ml IL-1b, 50 lM sulindac
enhanced significantly compared to the control as
sulfide, and/or 50 lM PDTC. After 3 h, TC1 levels were measured by
measured by semiquantitative RT-PCR . They are b-catenin
SYBR-green real-time PCR in quadruplicate using b-actin as internal
target genes and are implicated in cell proliferation (reviewed in
control. Sulindac and PDTC inhibited TC1 upregulation by IL-1bin a dose-dependent manner, suggesting an NF-jB-dependent
). Our data suggest that TC1 functions as a positive reg-
ulator of the Wnt/b-catenin signaling pathway in HK cells also.
Y. Kim et al. / FEBS Letters 580 (2006) 3519–3524
3.4. TC1 upregulates b-catenin target genes in HK cells
Previously, we have shown that TC1 upregulates b-catenin
target genes in cancer cells Because TC1 expression level
was quite low in HK line, a gain-of-function study by introduc-ing exogenous TC1 would have been appropriate. However, HK
cells are resistant to conventional gene transfer methods (data
not shown). Thus, we constructed the full length TC1 in lentiviral
vector to facilitate the gene transfer. HK cells were transfectedwith lentiviral constructs effectively so that most cells expressed
GFP in 24 h (data not shown). TC1-transfected HK cells showed
markedly upregulated TC1 expression in comparison with con-
trol vector-transfected cells as measured by real-time PCR
Fig. 2. Effect of sulindac and PDTC on IL-1b-induced TC1 upregu-
The expressions of cyclin D1, c-Myc, and c-Met were
lation. HK cells were treated with 20 U/ml IL-1b, 50 lM sulindac
enhanced significantly compared to the control as
sulfide, and/or 50 lM PDTC. After 3 h, TC1 levels were measured by
measured by semiquantitative RT-PCR . They are b-catenin
SYBR-green real-time PCR in quadruplicate using b-actin as internal
target genes and are implicated in cell proliferation (reviewed in
control. Sulindac and PDTC inhibited TC1 upregulation by IL-1bin a dose-dependent manner, suggesting an NF-jB-dependent
). Our data suggest that TC1 functions as a positive reg-
ulator of the Wnt/b-catenin signaling pathway in HK cells also.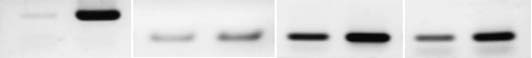 Y. Kim et al. / FEBS Letters 580 (2006) 3519–3524
Fig. 4. TC1 upregulates b-catenin target genes in HK cells. (A) TC1 expression in Lenti-TC1 or control vector-transfected cells. The expression wasmeasured by real-time PCR in quadruplicate using b-actin as internal control. (B) Semiquantitative RT-PCR for b-catenin target genes cyclin D1, c-Myc and c-Met in Lenti-TC1-transfected HK cells and control vector-transfected cells. Densitometric values were normalized using b-actin asloading control.
Y. Kim et al. / FEBS Letters 580 (2006) 3519–3524
Fig. 4. TC1 upregulates b-catenin target genes in HK cells. (A) TC1 expression in Lenti-TC1 or control vector-transfected cells. The expression wasmeasured by real-time PCR in quadruplicate using b-actin as internal control. (B) Semiquantitative RT-PCR for b-catenin target genes cyclin D1, c-Myc and c-Met in Lenti-TC1-transfected HK cells and control vector-transfected cells. Densitometric values were normalized using b-actin asloading control.