Do you want to buy antibiotics online without prescription? https://buyantibiotics24h.net/ - This is pharmacy online for you!
260857.qxd
Psychopharmacology (2001) 158:120–131DOI 10.1007/s002130100857
J. S. Rhodes · G. R. Hosack · I. Girard · A. E. Kelley G. S. Mitchell · T. Garland Jr
Differential sensitivity to acute administration of cocaine, GBR 12909, and fluoxetine in mice selectively bred for hyperactive wheel-running behavior
Received: 28 November 2000 / Accepted: 5 June 2001 / Published online: 9 August 2001 Springer-Verlag 2001
Abstract Rationale: To study the neural basis of genet-
tween genetically determined hyperactive wheel-run-
ic hyperactivity, we measured acute drug responses of
ning behavior and dysfunction in the dopaminergic neu-
mice (Mus domesticus) from four replicate lines that had
romodulatory system. Our selected lines may prove to
been selectively bred (23–24 generations) for increased
be a useful genetic model for attention deficit hyperac-
running-wheel activity. Objectives: We tested the hy-
pothesis that the high-running lines would respond dif-ferently to cocaine, GBR 12909, and fluoxetine (Prozac)
Keywords ADHD · Dopamine · Genetic selection ·
compared with four replicate, random-bred, control
Hyperactivity · Locomotor activity · Wheel running
lines. We also tested the hypothesis that the high-run-ning lines would display hyperactivity in cages withoutwheels. Methods: Drug trials were conducted at night,
during peak activity, after animals were habituated(3 weeks) to their cages with attached wheels. Revolu-
Understanding the genetic basis of behavior is one major
tions on wheels 10–40 min post-injection were used to
goal of neuroscience. Although genetic engineering con-
quantify drug responses. In a separate study, total photo-
tributes toward such an understanding (Xu et al. 1994;
beam breaks (produced on the first and second 24-h pe-
Baik et al. 1995; Giros et al. 1996), this approach has
riod of exposure) were used to quantify basal activity in
limitations. For example, if behavior is controlled by
animals deprived of wheels. Results: Cocaine and GBR
many genes working in concert, then the proportion of
12909 decreased wheel running in selected lines by re-
behavior explained by single-gene manipulations will be
ducing the average speed but not the duration of run-
small relative to the proportion explained by manipula-
ning, but these drugs had little effect in control lines.
tions that affect many genes (Smolen et al. 2000). Fur-
Fluoxetine reduced running speed and duration in both
thermore, if the behavioral effects of a single gene de-
selected and control animals, and the magnitude of the
pend on the genetic background, then genetically engi-
reduction was proportional to baseline activity. Basal
neered mice from inbred-strain progenitors may not ade-
activity in animals deprived of wheels (quantified using
quately represent similarly engineered mice from non-
photobeam breaks) was significantly higher in selec-
inbred populations (Crusio and Gerlai 1999; Cabib et al.
ted than control lines on the second day of testing. 2000). Conclusions: These results suggest an association be-
Artificial selection is a complementary tool to genetic
engineering studies of behavior, and it is well suited tothe study of complex traits controlled by many genes
J.S. Rhodes (✉) · G.R. Hosack · I. Girard · T. Garland Jr
Department of Zoology, University of Wisconsin,
(Garland and Carter 1994; Gibbs 1999). Selective-breed-
430 Lincoln Drive, Madison, WI 53706, USA
ing experiments have a long history in biology (Robertson
1980; Hill and Caballero 1992; Falconer and Mackay
Tel.: +1-608-2624437, Fax: +1-608-2656320
1996) and have been successfully employed in neurosci-
ence research (McClearn et al. 1978; Hausheer-Zarmakupi
Department of Psychiatry, School of Medicine,
et al. 1996; Marley et al. 1998). We used selective breed-
University of Wisconsin, 6001 Research Park Boulevard,
ing to increase voluntary wheel-running behavior in four
replicate lines derived from the same heterogeneous,
outbred base population of mice (Mus domesticus)
(Swallow et al. 1998; Koteja et al. 1999; Carter et al.
School of Veterinary Medicine, University of Wisconsin, 2015 Linden Drive, Madison, WI 53706, USA
2000; Rhodes et al. 2000; Bronikowski et al. 2001). Ge-
netic variation in the original base population (Hsd:ICR)
transporter (DAT) in mediating hyperactivity (Giros et
is similar to variation among individuals in wild popula-
al. 1996; Gainetdinov et al. 1999). Psychostimulant
tions of Mus domesticus (Rice and O’Brien 1980; Carter
drugs, such as cocaine and amphetamine, ameliorated
et al. 1999; and references therein).
hyperactivity in the DAT knockouts, similar to their ac-
After 17 generations of selective breeding, females
tions in ADHD subjects (Gainetdinov et al. 1999). Co-
(which we have chosen to study here) from our selected
caine primarily blocks DAT, but may secondarily block
lines displayed a 2.5-fold increase in the total number of
other amine reuptake transporters, such as SERT, the se-
revolutions run per day (Rhodes et al. 2000). Females
rotonin transporter (Womer et al. 1994). Gainetdinov et
from the selected lines have primarily increased their av-
al. (1999) suggested that cocaine attenuated the hyperac-
erage running speed rather than the amount of time spent
tivity exhibited by the DAT knockout mice through its
running (Swallow et al. 1998; Koteja et al. 1999; Rhodes
action on SERT, because the knockouts lacked DAT and
et al. 2000; Koteja and Garland 2001). The selected-line
fluoxetine (selective SERT inhibitor) caused a similar re-
females run in short bursts with short inter-bout pauses
duction in hyperactivity as did cocaine.
(Girard et al. 2001). As shown in this paper, the high-
We wished to test the acute effects of cocaine on hy-
running female mice also exhibit 24-h hyperactivity in
peractivity in our selected lines of mice and to determine
their cages (using photobeams to quantify activity) when
whether cocaine acted primarily through its actions on
they are deprived of wheels. Our selected lines of mice
SERT versus DAT. Therefore, after the cocaine trial, we
may therefore represent a novel murine model to study
conducted additional trials to measure the acute effects
the genetic basis of generalized 24-h hyperactivity, such
of fluoxetine and GBR 12909 (selective DAT inhibitor)
as that exhibited in human attention deficit hyperactivity
to evaluate the respective contributions of these neuro-
disorder (ADHD, Porrino et al. 1983).
Many workers have argued that genetic hyperactivity
in humans (and in the spontaneously hypertensive ratmodel of ADHD) is caused by impaired dopaminergic
function (Carey et al. 1998; Sagvolden and Sergeant1998; Papa et al. 2000; Russell 2000; Grace 2001; AnimalsSolanto et al. 2001), although ADHD has also been asso-
Mice from generations 23 and 24 of an artificial selection experi-
ciated with impaired noradrenergic function (Solanto
ment for high voluntary wheel-running behavior were studied (see
1998; Arnsten 2000, 2001). Hence, one plausible mecha-
Swallow et al. 1998 for details). The original progenitors were
nistic explanation for the increased activity in our select-
outbred, genetically variable (Rice and O’Brien 1980; Carter et al.
ed lines of mice is an alteration in dopaminergic func-
1999) laboratory house mice (Mus domesticus) of the Hsd:ICRstrain, purchased from Harlan Sprague Dawley in 1993. After two
tion. Pharmacological intervention can provide valuable
generations of random mating, mice were randomly paired and as-
insight as to whether a particular neurochemical system
signed to eight closed lines (ten pairs in each). In each subsequent
has been altered (Fink and Reis 1981; Cabib and Puglisi-
generation, when the offspring of these pairs were 6- to 8-weeks
Allegra 1985; Jones et al. 1991; Castner et al. 1993; old, they were housed individually with access to a running wheel
for 6 days, and a computer recorded wheel revolutions in 1-min
Giros et al. 1996; Giorgi et al. 1997; Henricks et al.
intervals [Wahman-type activity wheels (1.12-m circumference,
1997; Gainetdinov et al. 1999). For example, in the pres-
stainless steel and Plexiglas construction, Lafayette Instruments,
ent study, if hyperactive (selectively bred) animals re-
Lafayette, Ind.) were attached to standard clear plastic housing
sponded differently to drugs that affect dopaminergic
cages via a stainless-steel tube inserted into a hole in the wall ofthe cage]. In four “selected” lines, the highest-running (quantified
function, compared with control (unselected) animals,
as total number of revolutions run on day 5 and day 6 of the 6-day
then it could be inferred that some aspect of the dopami-
test) male and female from each family were chosen as breeders to
nergic system has been altered in the hyperactive ani-
propagate the lines to the next generation. In the four “control”
lines, a male and a female were randomly chosen from each fami-
We were interested in testing the effects of dopamine
ly. Within all lines, the chosen breeders were randomly paired ex-cept that sibling matings were not allowed.
reuptake inhibitors not only because dopamine has been
The purpose of having four replicate selected and four repli-
implicated in ADHD, but also because of the possible
cate control lines is to account for random genetic changes, such
roles that dopamine plays in motivation (Berridge and
as founder effects and genetic drift, which can cause lines to di-
Robinson 1998), reward (Di Chiara et al. 1993), and re-
verge even in the absence of selection. Any particular genetic orphenotypic difference between a given selected line and a given
inforcement (Damsma et al. 1992). Rodents are believed
random-bred control line may or may not be causally related to the
to perceive a reward from wheel running because it is
phenotype that was actually under selection. For example, in the
not a goal-oriented behavior and because they run volun-
present study, if we were to compare the drug responses of only
tarily (Sherwin 1998). Therefore, we hypothesized that
one hyperactive line with one control line, then we would have noway of determining whether any differences were the result of ran-
animals that exhibit increased wheel running may per-
dom genetic processes or the result of the selection for hyperactiv-
ceive altered incentive (Berridge and Robinson 1998) for
ity per se. Inferences about the causal factors underlying pheno-
typic changes in a selected line are greatly strengthened if repli-
Dopaminergic function has also been associated with
cate lines are maintained (Henderson 1989, 1997).
running speed (Freed and Yamamoto 1985) and locomo-
The Principles of Laboratory Animal Care (NIH publication
no. 85–23, revised 1985) was followed, and all experiments were
tion in general (Vallone et al. 2000). In particular, recent
approved by the University of Wisconsin Animal Care Committee.
“knockout” studies implicate the dopamine reuptake
Throughout the selection experiment and during this study, water
and food [Harlan Teklad Laboratory Rodent Diet (8604); aftergeneration 23, breeding females were given Harlan Teklad MouseBreeder Diet (7004)] were available ad libitum. Rooms were con-trolled for temperature (~22°C) and photoperiod 12-h/12-hlight/dark (lights on at 0700 hours, central standard time).
To simplify analyses, only females were used in the present
study. Different groups of mice were used for each of the drug tri-als that were conducted. For the cocaine trial, generation-23 ani-mals that were not among those chosen as breeders to propagatelines to the 24th generation were used. Because exclusion of thetop runners would have caused samples from the selected lines tobe biased downward with respect to wheel running, the lowest-running animals in selected-line families were also excluded. Ofthe remaining mice, 48 were randomly chosen to participate (sixper line, each from a different family).
To supply animals for the GBR 12909 and fluoxetine (Prozac)
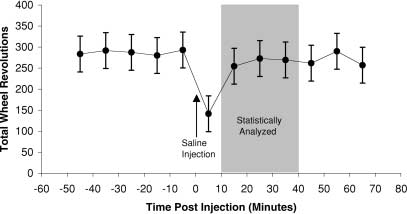
Fig. 1 The response of selected-line animals to the vehicle injec-
trials, generation-22 parents (that were not sacrificed for routine
tion during the cocaine trial. Data points represent least-square ad-
measurements) were allowed to produce a second litter. Six ani-
justed means ±SEM from a repeated-measures analysis of total
mals per line (from six separate families) were assigned to each of
wheel revolutions in 10-min increments. During the first 10-min
the GBR 12909 and fluoxetine trials. However, in line 1 (a ran-
period after injection, wheel running was substantially reduced.
dom-bred control line), only four litters were successfully weaned,
For this reason, we omitted the first 10 min in statistical analyses,
so only four animals were available for each trial. Similarly, in
line 8 (selected), only five animals were used in each trial. Thus,the experimental design for the fluoxetine and GBR trials wasslightly unbalanced.
suppressed by the vehicle injection during this period (see Fig. 1for an example). Within this 10- to 40-min period, total wheel rev-olutions, total number of minutes that the wheel showed at least
one revolution, and average speed of rotations (total number ofwheel rotations divided by number of minutes with any wheel rev-
The animals used for the cocaine trial were placed in cages with
access to running wheels in random order when they were approx-imately 68±1.4 days old (mean±SD). After 3 weeks of acclima-tion, mice were injected with either vehicle (0.9% saline) or co-
caine – 20 mg/kg or 40 mg/kg cocaine in a volume adjusted to thebody mass of the animal (0.01 ml/g). Animals were injected every
To determine whether the high wheel-running mice are also hyper-
other day for a total of three injection days, so that each individual
active when housed in cages without wheels, a separate group of
received all three types of injections (vehicle, medium, and high
32 female mice (4 per line) from generation 24 were used. Ani-
dose) over the course of the three injection days. Each mouse per
mals (not chosen as breeders; low-running individuals excluded
line received the three injections in a different order (one of the
by family, as described above for cocaine trials) were placed in the
six permutations of the three doses), randomized across lines, such
photobeam cages when they were approximately 57±2-days old.
that possible effects of injection order did not need to be consid-
Individual beam breaks (fine movements) and consecutive beam
breaks (coarse movements or ambulations) were recorded continu-
Mice were injected in random sequence, but the same se-
ously for 48 h using San Diego Instruments (San Diego, Calif.)
quence was used on each of the injection days. This was done so
software. Pine bedding, food, and water were available on the
that a mouse always received its injection at approximately the
floor of the cages. Rat-sized photobeam activity cages were used
same time of day. It usually took less than 2 min to capture, inject,
(dimensions 48×25×20 cm), and there was slight variation in the
and return a mouse to its home cage. Injections began 2 h after
distance of the photobeams from the floor of the cages. These dis-
lights off, during peak activity (unpublished data).
tances were measured and entered as covariates in the statistical
Animals for the GBR 12909 and fluoxetine trials were placed
analyses, along with body mass, because both these quantities
in cages with access to running wheels when they were approxi-
could affect the probability of photobeam breaks and consequently
mately 38±2.4 days old. Even though animals in the GBR 12909
obscure the actual relationship between cage activity and line type
and fluoxetine trials were younger than the mice in the cocaine
group, their running profiles were similar at the time they weregiven injections (see Results). Thus, we believe it is appropriate tocompare results across all three drug trials. Otherwise, the GBR
12909 and fluoxetine trials proceeded similarly to that for cocaine. Fluoxetine was administered at 10 mg/kg and 20 mg/kg, as was
SAS (SAS Institute Inc.) PROC MIXED (which employs restrict-
GBR 12909. Doses were chosen after consulting the literature ed maximum likelihood) was used to analyze the data. Line was(for cocaine, Iijima 1995; Giros et al. 1996; Marley et al. 1998;
always entered as a random effect nested within the fixed effect
Gainetdinov et al. 1999; for GBR 12909, Womer et al. 1994; line type (selected or control). The lines were separately propagat-Irifune et al. 1995; for fluoxetine, Possidente et al. 1992; Griebel
ed for 24 generations; thus, individuals in a given generation do
et al. 1995; Gainetdinov et al. 1999). In most of these studies,
not represent independent data points and must be nested within
however, drugs were administered during the day and wheel run-
the populations they arose from (Henderson 1989, 1997). Body
ning was not used to measure drug responses (but see Iijima
mass and wheel freeness (total number of revolutions produced by
1995). Therefore, we also conducted preliminary studies to deter-
the wheel after being accelerated to constant velocity, an inverse
mine behaviorally equivalent doses of the three uptake blockers.
measurement of how resistant the wheel is to continued rotation)
Wheel rotations were monitored via computer in 1-min inter-
were included as covariates in all the analyses of wheel-running
vals throughout each trial. We compared acute responses of select-
variables (except in those where wheel running variables were re-
ed and control animals (Womer et al. 1994; Marley et al. 1998;
gressed on each other). Stage of the estrus cycle was not measured
Gainetdinov et al. 1999), which we defined to be wheel running
and hence was not entered as a cofactor in any analyses.
produced in the 10- to 40-min period post-injection. The first
Baseline wheel running was compared between selected and
10 min was not included because wheel running was significantly
control lines by considering mean total revolutions during the
2 days preceding injections for all three drug trials combined. Data were analyzed using a two-way analysis of covariance(ANCOVA), including line type, drug trial, and the interaction be-tween drug trial and line type as cofactors.
To determine whether selected and control animals differen-
tially responded to the drugs, both the absolute and proportionalresponses were analyzed, because baseline wheel running differedbetween the selected and control lines (see Results). For the ab-solute response, the wheel-running variables (total revolutions,minutes with any revolutions, or average speed 10–40 min post-injection) were analyzed using repeated-measures two-factorANCOVA to test for an interaction between dose and line type (re-peated measures was needed to account for the fact that the threedoses were applied to the same individual on three separate days). Absolute responses were also analyzed separately for selected andcontrol lines to determine the effects of the drugs in each line type. For the separate analyses, a one-factor, repeated-measuresANCOVA was used to determine the effect of dose on the wheel-running variables. To improve normality of residuals, minuteswith any wheel rotations were always power transformed (see Y-axis legend of Fig. 3).
For the proportional response, a one-factor ANCOVA was
used to test for a line type effect on the ratio of the wheel-runningresponse after the high-dose injection to the response after the ve-hicle injection. To improve normality of residuals, the proportion-al responses were always rank transformed (data were highly posi-tively skewed otherwise).
To test for rate dependency of drug effects, linear regression
was used to determine the relationship between response to thehigh dose injection and baseline response to the vehicle injection. No covariates were entered in these analyses, because values foreach individual were regressed against each other.
For the cage activity data, a one-factor ANCOVA was used to
test for a line type effect on the total activity scores of the animals. Separate analyses were conducted for the first (novel) and second24 h of activity. Body mass and the distances of the photobeams tothe floor of the cages were always included as covariates. Fig. 2 A Mean wheel running (represented as total revolutions
Selective breeding for increased wheel running behavior
per day on day 5 and day 6 of a 6-day test; circumference of
has resulted in substantial divergence between the 4 se-
wheel = 1.12 m) of female mice from four replicate selected linesand four replicate control lines across generations. Wheel running
lected and 4 control lines in total number of revolutions
increased in each of the selected lines, but showed little change in
run per day (Fig. 2A). At generation 24 (see also Koteja
the control lines. B Mean number of minutes spent running (num-
and Garland 2001), female mice from selected lines
ber of 1-min intervals during which any revolutions were record-
(n=221) ran an average of 14,458 revolutions (16.2 km)
ed) for the same mice as in A. The time spent running did not di- verge substantially between selected and control lines. Female
per day (on day 5 and day 6 of the standard 6-day test),
mice in the selected lines accomplished more total revolutions per
representing a 2.78-fold increase over females from con-
day mainly by increasing their average running speed, rather than
trol lines (n=79), which ran an average of 5205 revolu-
tions (5.8 km) per day (Fig. 2A). The increase in wheelrunning was accomplished primarily by increased aver-age running speed (2.37-fold increase), because there
entered as cofactors, animals from all three drug trials
was only a 1.19-fold increase in the total number of min-
were considered simultaneously, n=137). Baseline level
utes with any revolutions (Fig. 2B).
of wheel running was similar for each drug trial (P value
As expected, the selected-line individuals used in for the effect of drug trial on mean revolutions 2 days
the three drug trials ran significantly more total revolu-
preceding injections = 0.63; P value for the interaction
tions than the control-line individuals. For example, con-
between drug trial and line type = 0.61).
sidering the mean total revolutions run on the 2 days preceding injections, animals from selected lines ran17,739±1032 versus 6946±1031 for control-line animals
(least-square adjusted means and standard errors from anested two-way ANCOVA, wheel freeness used as a co-
Dose–response profiles for cocaine and GBR 12909
variate, drug trial and drug trial by line type interaction
were strikingly similar (Fig. 3), suggesting that cocaine
Fig. 3A–I The wheel-running response to i.p. administration of
ANCOVA using data for selection-line animals only,
cocaine (left column), GBR 12909 (middle column), and fluoxe-
dose was a significant predictor of total revolutions
tine (right column) in mice from selected and control lines. Top
10–40 min post-injection (P<0.0001 for cocaine; P=0.001
row shows the total revolutions produced during the 10- to 40-minpost-injection period. Middle row shows the number of minutes
for GBR 12909). However, dose was not a significant
(power transformed to reflect the statistical analysis conducted)
predictor of total revolutions for control-line animals
with any wheel revolutions over the same time interval
(P=0.8203 for cocaine; P=0.2404 for GBR 12909).
(10–40 min post-injection). Bottom row shows the average speed
The dose-dependent decrease in total revolutions ob-
of running over the 10- to 40-min post-injection period. Dose–
served in selected animals in response to the DAT inhibi-
response profiles are similar for GBR 12909 and cocaine, but dif-ferent for fluoxetine. Cocaine and GBR 12909 attenuated the total
tors was caused by a decrease in the speed of running,
revolutions by reducing the speed, not the number of active min-
not by a decrease in the number of minutes active in the
utes in mice from selected lines. Total revolutions for control ani-
wheel (Fig. 3D, E, G, H). Dose did not affect the number
mals remained the same because the number of minutes spent run-
of minutes spent running 10–40 minutes post-injection in
ning increased, whereas speed slightly decreased. Fluoxetine, incontrast, reduced the total, speed, and minutes of revolutions in
selection animals (P=0.22 for cocaine, and P=0.67 for
both selected and control animals. Least-square adjusted means
GBR 12909, one-way ANCOVA). The minutes variable
and standard error bars are shown. P values for interactions be-
was negatively skewed and was raised to the tenth power
tween dose and line type using a two-way repeated-measures anal-
so that residuals were approximately normally distribut-
ed. Figure 3 reports the least-square means for the trans-formed minutes variable to reflect the statistical analyses
acted by blocking DAT. Selected and control animals re-
that were conducted. The untransformed means for min-
sponded differently to cocaine whether or not the re-
utes of wheel running in selected-line animals were 24.7,
sponse was measured on an absolute scale or as a pro-
22.3, and 23.6 for cocaine doses 0, 20, and 40 mg/kg,
portion of the baseline response to the vehicle injection
and 26.9, 27.5, and 26.9 for GBR 12909 doses 0, 10, and
(Table 1). Statistical results were similar for GBR 12909,
although the effect of line type on proportional responses
Total revolutions in control animals did not change in
response to injection of cocaine or GBR 12909 (Fig. 3A,
Cocaine and GBR 12909 dose dependently decreased
B), because speed slightly decreased while number of
total revolutions run during the 10- to 40-min period
minutes increased (Fig. 3D, E, G, H). Dose was a signifi-
post-injection in selected-line animals but had little ef-
cant predictor of minutes of wheel running in control-
fect in control-line animals (Fig. 3A, B). In a one-factor
line animals (P=0.0003 for cocaine, P<0.0001 for GBR
Table 1 Analysis of variance (ANOVA) table for statistical ana-
sponses were rank transformed and LS adjusted means ± SEM of
lyses of the drug response data. The P values in the left-hand col-
the ranks are shown for control and selected lines. Higher rank in-
umn indicate differences in absolute responses to the drugs [least-
dicates reduced sensitivity to the drug. Numerator degrees of free-
square (LS) means for these analyses are displayed graphically in
dom (NDF), denominator degrees of freedom (DDF), and F statis-
Fig. 3]. The P values in the right-hand column indicate differences
tics are also presented. P values less than 0.05 are in bold. Body
in proportional responses to the drugs. The proportional response
mass and wheel freeness (an inverse measure of wheel resistance)
was quantified as the response after the high-dose injection divid-
were included as covariates in all analyses and were occasionally
ed by the response after the vehicle injection. Proportional re-
significant. Sample size = 45–48 individuals for each analysis
One-factor analysis of covariance for line type
effect on rank-transformed proportional response
<0.0001 <0.0001 <0.0001
12909, one-factor ANCOVA). The untransformed means
However, the proportional decrease in wheel running af-
for minutes of wheel-running activity in control-line ani-
ter fluoxetine administration was similar for mice from
mals were 22.9, 25.9, and 28.4 for cocaine doses 0, 20,
selected and control lines (Table 1).
and 40 mg/kg, and 21.9, 27, and 29.5 for GBR 12909
In each of the drug trials, individual response to the
vehicle injection was a significant linear predictor of re-
The fact that the DAT inhibitors did not increase min-
sponse to the high dose injection (Fig. 4, P value for the
utes of running in selected animals (but did in controls)
slope of the linear regression <0.0001 for cocaine,
is not a consequence of a ceiling effect, but rather is evi-
P=0.007 for GBR 12909, and P=0.0003 for fluoxetine;
dence that the selected and control animals responded
line was entered as a random effect, but line type and the
differently to these drugs. The number of minutes spent
interaction between line type and vehicle injection were
running during the 10- to 40-min period after the vehicle
not significant and, hence, were removed from the mod-
injection was similar in selected and control animals for
el; no covariates were entered). The slope was positive
mice used in the cocaine and GBR 12909 trials (one-way
but less than unity in each case (Fig. 4). The intercept
ANCOVA effect of line type on the vehicle response:
was significantly positive for cocaine (P=0.0031) and
P=0.48 for cocaine and P=0.11 for GBR 12909). Further,
GBR 12909 (P=0.0041) but was not significantly differ-
the greatest number of minutes of running occurred in
ent from 0 for fluoxetine (P=0.25). Thus, it appears that
control-line animals given the high doses of DAT inhibi-
the DAT inhibitors increased wheel running in individu-
tors (see raw values above or Fig. 3 for transformed als with low baseline levels and decreased wheel runningvalues).
in individuals with relatively high baseline levels. In the
In contrast to results for GBR 12909 and cocaine, flu-
previous analyses, we did not detect an effect of the DAT
oxetine decreased total revolutions, speed, and time
inhibitors on mean total revolutions among control-line
spent running in both selected and control animals
animals because approximately half the control-line ani-
(Fig. 3F, I). Dose was a significant predictor of total rev-
mals were stimulated and half were suppressed by the
olutions, speed, and minutes of wheel running in control-
line animals (P=0.004 for total, P<0.002 for speed,
Wheel freeness and body mass were only occasion-
P=0.05 for minutes, one-factor ANCOVA) and in select-
ally significant predictors of wheel-running responses.
ed-line animals (P<0.0001 for total, P<0.0001 for min-
When they were significant, wheel freeness was posi-
utes, P<0.0001 for speed). The untransformed means for
tively related to wheel running, and body mass was
minutes of wheel running were 23, 17.9, and 16.2 for
negatively related to wheel running. The random effect
control-line animals given fluoxetine doses 0, 10, and
of line nested within line type was not significant in
20 mg/kg, respectively, and 28.4, 26.1, and 19.3 for se-
any analyses, indicating that genetic drift or founding
lected-line animals. The absolute decrease in total revo-
effects had not significantly altered the traits that were
lutions, time running, and average speed was greater in
measured here. As expected (for example, see Swallow
magnitude for selected-line animals than controls, and
et al. 1999), control-line animals were significantly
the interaction between dose and line type in the two-
heavier in body mass in all data sets except that for
factor ANCOVA was statistically significant (Table 1).
ambulation; P=0.0004; Fig. 5) during the second 24-hperiod of testing. However, there were no significant dif-ferences on the first day of testing (P=0.26 for finemovements and P=0.07 for coarse movements; Fig. 5). On the first day, both selected and control animals ex-hibited relatively high levels of spontaneous activity(Fig. 5). By the second day, control animals displayedlower levels of activity, while selected animals continuedto display high levels. In addition to the line type effects,both the distance of the photobeams from the floor of thecage and body mass were significant predictors of cageactivity, as measured by the photobeam breaks. Distancewas negatively related to photobeam counts, and bodymass was positively related.
We have developed a new animal model to study genetichyperactivity: lines of mice artificially selected for in-creased voluntary wheel-running behavior (Swallow et al. 1998, 1999; Koteja et al. 1999; Carter et al. 2000; Rhodeset al. 2000; Koteja and Garland 2001; Bronikowski et al. 2001). In this study, we found that our high-running miceare also hyperactive in their cages when deprived ofwheels, as demonstrated using photobeams to measureactivity (Fig. 5). We also found that control and selectedanimals responded differently to drugs that inhibit the do-pamine transporter protein. Cocaine and GBR 12909 re-duced wheel running in hyperactive animals, but thesesame drugs had no average effect in control-line animals. The ability to partition total wheel revolutions into min-utes of revolutions and average speed enabled us to showthat the reduction in total wheel revolutions (by cocaineand GBR 12909) in selected lines was caused by a reduc-tion in the average speed of running, not the number ofminutes spent running. This result is particularly impor-tant because it shows that cocaine and GBR 12909 ame-
Fig. 4 Individual responses (total revolutions 10–40 min post-
liorated the hyperactivity as it is normally expressed by
injection) after the high-dose injection plotted against responses
our selected-line females, which is mainly by increased
following vehicle injection for cocaine (top), GBR 12909 (middle),
speed of wheel running (see Fig. 2 and above references).
and fluoxetine (bottom). The line of unity (dashed) is drawn toshow the extent and direction of drug effects for each individualmouse. Also drawn are the regression lines (solid) to show the av-erage response predicted by baseline rates of activity. Cocaine and
GBR 12909 tended to stimulate wheel running in individuals withlow baseline rates, but depressed activity in individuals with high
The simplest neurochemical explanation of our results is
baseline rates (intercept of the regression line was significantly
that dopaminergic function is altered in the selection
positive). In contrast, fluoxetine depressed wheel running irrespec-tive of baseline response (intercept of the regression line was not
lines. We reached this conclusion because the DAT in-
hibitors elicited a proportionately greater response in se-lected than control lines. However, a more complicatedinterpretation is also possible. In theory, an alteration in
any neurochemical system pre- or post-synaptically asso-ciated with dopaminergic neurons could have influenced
Mice from selected lines exhibited higher numbers of
the response to the DAT inhibitors. Clearly, further re-
both fine movements (counts of individual photobeam
search is needed to fully characterize the neurochemical
breaks; nested ANCOVA P=0.048) and coarse move-
alterations in the selectively bred, hyperactive lines of
ments (counts of consecutive beam breaks, also termed
mice. However, the present results are important because
Fig. 5 Activity of mice from selected and control lines as re- corded with photobeams over a 48-h period using rat-size cages (dimensions 48×25×20 cm). The left column represents the first 24 h; right column the second 24 h. The top row shows fine movements; bottom row coarse movements or “am- bulations”. P values are from nested analysis of covariance models, with line nested within line type, and such covariates as body mass. During the initial 24-h period, no significant dif- ferences were observed for fine or coarse movements; but, dur- ing the second day, mice from selected lines exhibited many more ambulations or coarse movements than controls, and moderately more fine move- ments. Least-square adjusted means and standard error bars are shown
they provide an a priori hypothesis to test: reduced dopa-
1988). However, before diffusing to extrasynaptic spaces,
minergic function is associated with genetic hyperactivi-
dopamine would stimulate postsynaptic receptors, and the
ty in our mice. Recent evidence suggests that dopaminer-
time course for diffusion to extrasynaptic spaces is un-
gic systems modulate incentive to acquire a reward, not
the hedonic impact of the reward itself (see Berridge and
In this study, we utilized acute rather than chronic
Robinson 1998 for a review of the incentive salience hy-
drug administration to investigate the involvement of do-
pothesis for the role of dopamine in reward). Therefore,
pamine and serotonin in the hyperactive wheel running.
an alteration in dopaminergic function is a reasonable
Therapeutic efficacy of DAT inhibitors occurs immedi-
mechanism for the hyperactive running, presuming that
ately (within 2 h of oral administration, Solanto 1998)
the mice perceive a reward from wheel running (Sherwin
and is not thought to involve long-term adaptive re-
sponses (Solanto et al. 2001). However, SERT inhibitorsoften require chronic administration over days or evenweeks. The mechanisms responsible for behavioral alter-
ation after chronic exposure are complicated by suchprocesses as receptor downregulation, induction of neu-
Genetic hyperactivity in humans is believed to result from
rotrophins, and even neurogenesis (Jacobs et al. 2000;
abnormally low tonic dopaminergic activity within the nu-
Stamford et al. 2000). To avoid these unintended compli-
cleus accumbens, leading to abnormally highphasic dopa-
cations, we felt it was essential to restrict our initial in-
mine responses (Grace 2001; ADHD has also been associ-
ated with altered noradrenergic function, Solanto 1998,2001; Arnsten 2000, 2001). Results of our drug trials areconsistent with this dopamine hypothesis, if one assumes
that cocaine and GBR 12909 ameliorated the hyperactivi-ty in our selected lines as a consequence of drug-induced
Rate-dependent effects of drugs occur when the drug ef-
increases in dopamine concentrations in extrasynaptic
fect is related to the control rate of response (Robbins
spaces within the nucleus accumbens. Increased dopamine
and Sahakian 1979). An inverse relationship between
in extrasynaptic spaces would increase stimulation of au-
control rate and drug effect is generally found after treat-
toreceptors, which would downregulate the spike-depen-
ment with DAT inhibitors (Sanger and Blackman 1976;
dent phasic component of dopamine release (Grace 2001).
Robbins and Sahakian 1979). Our results are consistent
Cocaine is known preferentially to increase dopamine
with an inverse rate-dependent effect for cocaine and
concentrations in the accumbens (Di Chiara and Imperato
GBR 12909, because individuals with low basal activity
scores tended to be aroused by the drugs, whereas indi-
Gainetdinov et al. (1999) concluded that cocaine calmed
viduals with high basal scores were depressed (Fig. 4).
the DAT knockouts through its actions on the serotonin
This does not contradict the result that average total rev-
system, whereas cocaine appeared to act on the dopami-
olutions in control-line animals did not change in re-
nergic system in our lines (based on comparison with re-
sponse to the DAT inhibitors because approximately half
sults of GBR 12909 trials). Thus, comparison of the
the control-line animals were stimulated and half were
DAT knockout mice with our high wheel-running mice
suppressed by the drugs (see Results, Fig. 4). Similarly,
suggests that hyperactivity may come in different forms
reports that normal and hyperactive humans respond
with potentially different underlying mechanisms.
qualitatively similarly to therapeutic doses of methylphe-
A selection experiment for open-field behavior
nidate (Ritalin) and D-amphetamine (Rapoport et al.
(DeFries et al. 1970) and interspecies comparative data
1978; Aman et al. 1984; Solanto 1998, Solanto et al.
provide further evidence that hyperactivity in the habitu-
2001) is not inconsistent with inverse rate dependency
ated versus novel environment is controlled by different
(Robbins and Sahakian 1979). As pointed out by Millard
underlying mechanisms. Lines of mice selected for in-
and Standish (1982), the mechanistic explanation for in-
creased activity in an open-field arena did not exhibit in-
verse rate dependency is not known. Thus, rate depen-
creased spontaneous wheel-running activity (DeFries et
dency is not a suitable explanation for the “paradoxical”
al. 1970) and our high-running lines (which also display
effect that stimulants have on individuals at either end of
hyperactivity in photobeam cages after 24 h of acclima-
the activity continuum, but rather is a description of an
tion) are not hyperactive in the open-field arena
empirical observation (Millard and Standish 1982).
(Bronikowski et al. 2001). Further, as noted by
The fact that cocaine and GBR 12909 did not produce
Bronikowski et al. (2001), across 12 species of muroid
an average increase in total wheel revolutions in the rodents, the correlation between open-field activity andcontrol-line animals is not surprising because the drug
wheel running is not significantly different from zero.
trials were conducted at night, during peak activity (see
Thus, all studies to date support the view that spontane-
Methods). Typically, drug trials are conducted during the
ous activity in a habituated environment and locomotor
day when nocturnal rodents are normally sleeping (Reith
behavior in a novel open-field environment are not con-
1986; George 1989; Ichihara et al. 1993; Womer et al.
trolled by the same underlying mechanisms.
1994; Irifune et al. 1995). During the day, cocaine and
What form of hyperactivity is exhibited by people di-
GBR 12909 are known to stimulate activity in rodents
agnosed as having ADHD? According to Porrino et al.
(Kelley et al. 1989; Gainetdinov et al. 1999). However,
(1983), ADHD children exhibit 24-h hyperactivity, in-
baseline levels of activity are near zero during the day
cluding during sleep. However, ADHD children may not
(Gainetdinov et al. 1999), and a floor effect limits the
exhibit hyperactivity in the novel or stressful environ-
possible direction of response. At night, mice could re-
ment, such as during an experimental trial or at the doc-
spond by either increasing or decreasing activity levels.
tor’s office (Sleator and Ullmann 1981; Sagvolden and
For example, in male ddY mice, 40 mg/kg cocaine sup-
Sergeant 1998). Hyperactivity in humans is primarily
pressed night-time wheel running (Iijima et al. 1995). treated with Ritalin and D-amphetamine, drugs which actInterestingly, male ddY mice exhibit relatively high more similarly to cocaine and GBR 12909 than to fluox-levels of spontaneous wheel running [8.7 km/day, esti-
etine. However, fluoxetine (Prozac) is occasionally given
mated from raw data from Iijima et al. (1995) versus
to ameliorate hyperactivity in humans (Barrickman et al.
5.8 km/day in our control-line females and 5.0 km/day in
1991). We are currently conducting drug trials with Ri-
our control-line males (data from generation 24, as pre-
talin to further validate the selected lines as a model for
sented in Koteja and Garland 2001)].
With respect to implications for ADHD, one unusual
feature of the present experiments is that we studied fe-
Behavioral profiles of hyperactive animals
male mice, even though hyperactivity is two- to ninefoldmore prevalent in male children (Andersen and Teicher
It is useful to compare the behavior of our selected-line
et al. 2000; but note that ADHD may be more similarly
animals with the DAT knockout mice because both are
represented in male and female adults, and some argue
hyperactive. If behavioral profiles are similar, then we
that ADHD may be over-diagnosed in male children rel-
might infer that the hyperactivity in our selected lines is
ative to female children, Biederman et al. 1994). We de-
caused by impaired DAT. However, behavioral profiles
cided to study females to make use of the fact that select-
are not similar. First, the difference in activity between
ed-line females increase their total number of revolutions
the DAT knockouts and the wild-type controls decreased
primarily by increasing the speed of running, not the
with trial duration in a 3-h test using photobeam activity
number of minutes spent active, whereas males show a
cages (217×268×104 mm), such that hyperactivity in the
greater increase in duration of activity, although they too
DAT knockouts was most apparent in the novel environ-
mainly show increased speed (Swallow et al. 1998,
ment (at the beginning of the experimental trial; Giros et
1999; Koteja et al. 1999; Rhodes et al. 2000; Koteja and
al. 1996). In contrast, hyperactivity in our selected lines
Garland 2001). Also, female mice generally run more to-
was most apparent in the habituated environment (on tal revolutions than males (previous references), whichthe second day of photobeam testing; Fig. 5). Second,
may enhance the ability to detect drug effects.
Arnsten AF (2000) Genetics of childhood disorders: XVIII.
ADHD, Part. 2: norepinephrine has a critical modulatory in-fluence on prefrontal cortical function. J Am Acad Child
Both pharmacokinetic (rate of absorption, distribution or
excretion of drugs) and pharmacodynamic (neurochemi-
Arnsten AFT (2001) Dopaminergic and noradrenergic influences
cal) variables could have contributed to the divergent re-
on cognitive function mediated by prefrontal cortex. In:
sponses of animals from our selected and control lines
Solanto MV, Arnsten AFT, Castellanos FX (eds) Stimulant
(Benuck et al. 1987; Henricks et al. 1997; Wiener and
drugs and ADHD. Oxford University Press, Oxford, pp 185–208
Reith 1990). For example, one trait that has diverged be-
Baik JH, Picetti R, Saiardi A, Thiriet G, Dierich A, Depaulis A,
tween selected and control lines, and which might affect
Le Meur M, Borrelli E (1995) Parkinsonian-like locomotor
pharmacokinetics, is body mass (hyperactive animals are
impairment in mice lacking dopamine D2 receptors. Nature
smaller: Swallow et al. 1999). However, all statistical an-
Barrickman L, Noyes R, Kuperman S, Schumacher E, Verda M
alyses were conducted with body mass as a covariate,
(1991) Treatment of ADHD with fluoxetine: a preliminary
thus effectively removing its influence from group com-
trial. J Am Acad Child Adolesc Psychiatry 30:762–767
Benuck M, Lajtha A, Reith ME (1987) Pharmacokinetics of sys-
Evidence from the literature also argues against a
temically administered cocaine and locomotor stimulation inmice. J Pharmacol Exp Ther 243:144–149
pharmacokinetic explanation of divergent drug responses
Berridge CW, Mitton E, Clark W, Roth RH (1999) Engagement in
among animals that differ in baseline activity (Benuck
a non-escape (displacement) behavior elicits a selective and
et al. 1987). Benuck et al. (1987) reported no correlation
lateralized suppression of frontal cortical dopaminergic utiliza-
between baseline activity and brain cocaine concentra-
tions measured 12, 22, and 32 min after i.p. injections of
Berridge KC, Robinson TE (1998) What is the role of dopamine in
reward: hedonic impact, reward learning, or incentive sa-
25 mg/kg cocaine in the BALB inbred mouse strain.
lience? Brain Res Brain Res Rev 28:309–369
These results may not be relevant if the physiological ba-
Biederman J, Faraone SV, Spencer T, Wilens T, Mick E, Lapey
sis for the variation in baseline activity differs in the
KA (1994) Genetic differences in a sample of adults with at-
BALB mice relative to our genetic lines. The possibility
tention deficit hyperactivity disorder. Psychiatry Res 53:13–29
Bronikowski AM, Carter PA, Swallow JG, Girard IA, Rhodes JS,
that pharmacokinetic differences contributed to the diver-
Garland T Jr (2001) Open-field behavior of house mice artifi-
gent responses reported here cannot be ruled out entirely.
cially selected for high voluntary wheel running. Behav Genet(in press)
Cabib S, Puglisi-Allegra S (1985) Different effects of apomor-
phine on climbing behavior and locomotor activity in three
strains of mice. Pharmacol Biochem Behav 23:555–557
Cabib S, Orsini C, Le Moal M, Piazza PV (2000) Abolition and
Results presented here are consistent with the hypothesis
reversal of strain differences in behavioral responses to drugs
that genetic hyperactivity is associated with altered do-
of abuse after a brief experience. Science 289:463–465
paminergic function (Carey et al. 1998; Sagvolden and
Carey MP, Diewald LM, Esposito FJ, Pellicano MP, Gironi Carne-
vale UA, Sergeant JA, Papa M, Sadile AG (1998) Differential
Sergeant 1998; Papa et al. 2000; Russell 2000; Grace
distribution, affinity and plasticity of dopamine D-1 and D-2
2001; Solanto 2001). In future studies, we will quantify
receptors in the target sites of the mesolimbic system in an an-
dopaminergic function using microdialysis (Damsma et
imal model of ADHD. Behav Brain Res 94:173–185
al. 1992; Meeusen and De Meirleir 1995) or by measur-
Carter PA, Garland T Jr, Dohm MR, Hayes JP (1999) Genetic vari-
ation and correlations between genotype and locomotor physi-
ing dopamine and the primary dopamine catabolite
ology in outbred laboratory house mice (Mus domesticus).
(DOPAC) concentrations in micro-dissected brain re-
gions using high-performance liquid chromatography
Carter PA, Swallow JG, Davis SJ, Garland T Jr (2000) Nesting be-
(Berridge et al. 1999). Pharmacological manipulation
havior of house mice (Mus domesticus) selected for increasedwheel-running activity. Behav Genet 30:85–94
combined with direct measurement of concentrations of
Castner SA, Xiao L, Becker JB (1993) Sex differences in striatal
neurochemicals in regional areas of the brain are power-
dopamine: in vivo microdialysis and behavioral studies. Brain
ful tools; when applied to our replicate, selectively bred,
hyperactive lines of mice, strong inference regarding the
Crusio WE, Gerlai RT (1999) Behavioral neurogenetics: the new
neural basis of genetic hyperactivity will be possible.
synthesis. In: Crusio WE, Gerlai RT (eds) Handbook of mo-lecular-genetic techniques for brain and behavior research. Techniques in the behavioral and neural sciences, vol 13. Else-
Acknowledgements Supported in part by NSF grant IBN-9728434
to T.G., NIH HL 36780 and 53319 to G.S.M., and NIH grant P30
Damsma G, Pfaus JG, Wenkstern D, Phillips AG, Fibiger HC
HD03352 (P.I.: Terrence R. Dolan). The cocaine was supplied to
(1992) Sexual behavior increases dopamine transmission in
A.E.K. by the National Institute on Drug Abuse.
the nucleus accumbens and striatum of male rats: comparisonwith novelty and locomotion. Behav Neurosci 106:181–191
DeFries JC, Wilson JR, McClearn GE (1970) Open-field behavior
in mice: selection response and situational generality. Behav
Di Chiara G, Imperato A (1988) Drugs abused by humans prefer-
Aman MG, Vamos M, Werry JS (1984) Effects of methylpheni-
entially increase synaptic dopamine concentrations in the me-
date in normal adults with reference to drug action in hyperac-
solimbic system of freely moving rats. Proc Natl Acad Sci
Andersen SL, Teicher MH (2000) Sex differences in dopamine re-
Di Chiara G, Acquas E, Tanda G, Cadoni C (1993) Drugs of
ceptors and their relevance to ADHD. Neurosci Biobehav Rev
abuse: biochemical surrogates of specific aspects of natural re-
Falconer DS, Mackay TFC (1996) Introduction to quantitative ge-
Koteja P, Garland T Jr (2001) Response to R. Eikelboom. Anim
Fink JS, Reis DJ (1981) Genetic variations in midbrain dopamine
Koteja P, Garland T Jr, Sax JK, Swallow JG, Carter PA (1999) Be-
cell number: parallel with differences in responses to dopami-
haviour of house mice artificially selected for high levels of
nergic agonists and in naturalistic behaviors mediated by cen-
voluntary wheel running. Anim Behav 58:1307–1318
tral dopaminergic systems. Brain Res 222:335–349
McClearn GE, Kakihana R (1978) Selective breeding for ethanol
Freed CR, Yamamoto BK (1985) Regional brain dopamine metab-
sensitivity: short-sleep and long-sleep mice. In: McClearn GE,
olism: a marker for the speed, direction, and posture of mov-
Deitrich RA, Erwin VG (eds) Development of animal models
as pharmacogenetic tools: proceedings of a workshop. US De-
Gainetdinov RR, Wetsel WC, Jones SR, Levin ED, Jaber M,
partment of Health and Human Services Public Health Service
Caron MG (1999) Role of serotonin in the paradoxical calm-
Alcohol Drug Abuse and Mental Health Administration – Na-
ing effect of psychostimulants on hyperactivity. Science 283:
tional Institute on Alcohol Abuse and Alcoholism. Research
Monograph No. 6., 4–6 December 1978, pp 147–159
Garland T Jr, Carter PA (1994) Evolutionary physiology. Annu
Marley RJ, Arros DM, Henricks KK, Marley ME, Miner LL
(1998) Sensitivity to cocaine and amphetamine among mice
George FR (1989) Cocaine produces low dose locomotor depres-
selectively bred for differential cocaine sensitivity. Psycho-
sant effects in mice. Psychopharmacology 99:147–150
Gibbs AG (1999) Laboratory selection for the comparative physi-
Meeusen R, De Meirleir K (1995) Exercise and brain neurotrans-
Giorgi O, Corda MG, Carboni G, Frau V, Valentini V, Di Chiara G
Millard WJ, Standish LJ (1982) The paradoxical effect of central
(1997) Effects of cocaine and morphine in rats from two psy-
nervous system stimulants on hyperactivity: a paradox unex-
chogenetically selected lines: a behavioral and brain dialysis
plained by the rate-dependent effect. J Nerv Ment Dis
Girard I, McAleer MA, Rhodes JS, Garland T Jr (2000) Increased
Papa M, Sellitti S, Sadile AG (2000) Remodeling of neural net-
intermittency of locomotion in house mice selectively bred for
works in the anterior forebrain of an animal model of hyperac-
high voluntary wheel running (abstract). Am Zool 40:1029
tivity and attention deficits as monitored by molecular imag-
Giros B, Jaber M, Jones SR, Wightman RM, Caron MG (1996)
ing probes. Neurosci Biobehav Rev 24:149–156
Hyperlocomotion and indifference to cocaine and amphet-
Porrino LJ, Rapoport JL, Behar D, Sceery W, Ismond DR, Bunney
amine in mice lacking the dopamine transporter. Nature
WE Jr (1983) A naturalistic assessment of the motor activity
of hyperactive boys. I. Comparison with normal controls. Arch
Grace AA (2001) Psychostimulant actions on dopamine and lim-
bic system function: relevance to the pathophysiology and
Possidente B, Lumia AR, McEldowney S, Rapp M (1992) Fluoxe-
treatment of ADHD. In: Solanto MV, Arnsten AFT, Castellanos
tine shortens circadian period for wheel running activity in
FX (eds) Stimulant drugs and ADHD. Oxford University
Rapoport JL, Buchsbaum MS, Zahn TP, Weingartner H, Ludlow
Griebel G, Blanchard DC, Agnes RS, Blanchard RJ (1995) Differ-
C, Mikkelsen EJ (1978) Dextroamphetamine: cognitive and
ential modulation of antipredator defensive behavior in Swiss-
behavioral effects in normal prepubertal boys. Science 199:
Webster mice following acute or chronic administration of
imipramine and fluoxetine. Psychopharmacology 120:57–66
Reith ME (1986) Effect of repeated administration of various dos-
Hausheer-Zarmakupi Z, Wolfer DP, Leisinger-Trigona MC, Lipp
es of cocaine and WIN 35,065–2 on locomotor behavior of
HP (1996) Selective breeding for extremes in open-field activ-
ity of mice entails a differentiation of hippocampal mossy fi-
Rhodes JS, Koteja P, Swallow JG, Carter PA, Garland T Jr (2000)
Body temperatures of house mice artificially selected for high
Henderson ND (1989) Interpreting studies that compare high- and
voluntary wheel-running behavior: repeatability and effect of
low-selected lines on new characters. Behav Genet 19:473–
genetic selection. J Therm Biol 25:391–400
Rice MC, O’Brien SJ (1980) Genetic variance of laboratory out-
Henderson ND (1997) Spurious associations in unreplicated se-
Robbins TW, Sahakian BJ (1979) “Paradoxical” effects of psycho-
Henricks KK, Miner LL, Marley RJ (1997) Differential cocaine
motor stimulant drugs in hyperactive children from the stand-
sensitivity between two closely related substrains of C57BL
point of behavioural pharmacology. Neuropharmacology 18:
Hill WG, Caballero A (1992) Artificial selection experiments.
Robertson A (ed) (1980) Selection experiments in laboratory
and domestic animals. Commonwealth Agricultural Bureau,
Ichihara K, Nabeshima T, Kameyama T (1993) Mediation of do-
pamine D1 and D2 receptors in the effects of GBR 12909 on
Russell VA (2000) The nucleus accumbens motor-limbic interface
latent learning and locomotor activity in mice. Eur J Pharma-
of the spontaneously hypertensive rat as studied in vitro by the
superfusion slice technique. Neurosci Biobehav Rev 24:133–
Iijima Y, Shinoda M, Kuribara H, Asami T, Uchihashi Y (1995)
Evaluation of acute and sub-acute effects of cocaine by means
Sagvolden T, Sergeant JA (1998) Attention deficit/hyperactivity
of circadian variation in wheel-running and drinking in mice.
disorder – from brain dysfunctions to behaviour. Behav Brain
Nihon Shinkei Seishin Yakurigaku Zasshi 15:315–321
Irifune M, Nomoto M, Fukuda T (1995) Effects of GBR 12909 on
Sanger DJ, Blackman DE (1976) Rate-dependent effects of drugs:
locomotor activity and dopamine turnover in mice: compari-
a review of the literature. Pharmacol Biochem Behav 4:73–83
son with apomorphine. Eur J Pharmacol 272:79–85
Sherwin CM (1998) Voluntary wheel running: a review and novel
Jacobs BL, Praag H, Gage FH (2000) Adult brain neurogenesis
and psychiatry: a novel theory of depression. Mol Psychiatry
Sleator EK, Ullmann RK (1981) Can the physician diagnose hy-
peractivity in the office? Pediatrics 67:13–17
Jones BC, Campbell AD, Radcliffe RA, Erwin VG (1991) Co-
Smolen P, Baxter DA, Byrne JH (2000) Mathematical modeling of
caine actions, brain levels and receptors in selected lines of
mice. Pharmacol Biochem Behav 40:941–948
Solanto MV (1998) Neuropsychopharmacological mechanisms
Kelley AE, Lang CG (1989) Effects of GBR 12909, a selective
of stimulant drug action in attention-deficit hyperactivity dis-
dopamine uptake inhibitor, on motor activity and operant be-
order: a review and integration. Behav Brain Res 94:127–
havior in the rat. Eur J Pharmacol 167:385–395
Solanto MV, Arnsten AFT, Castellanos FX (2001) The neurosci-
Vallone D, Picetti R, Borrelli E (2000) Structure and function of
ence of stimulant drug action in ADHD. In: Solanto MV,
dopamine receptors. Neurosci Biobehav Rev 24:125–32
Arnsten AFT, Castellanos FX (eds) Stimulant drugs and
Wiener HL, Reith ME (1990) Correlation between cocaine-in-
ADHD. Oxford University Press, Oxford, pp 185–208
duced locomotion and cocaine disposition in the brain among
Stamford JA, Davidson C, McLaughlin DP, Hopwood SE (2000)
four inbred strains of mice. Pharmacol Biochem Behav
Control of dorsal raphe 5-HT function by multiple 5-HT(1)
autoreceptors: parallel purposes or pointless plurality? Trends
Womer DE, Jones BC, Erwin VG (1994) Characterization of do-
pamine transporter and locomotor effects of cocaine, GBR
Swallow JG, Carter PA, Garland T Jr (1998) Artificial selection
12909, epidepride, and SCH 23390 in C57BL and DBA mice.
for increased wheel-running behavior in house mice. Behav
Xu M, Moratalla R, Gold LH, Hiroi N, Koob GF, Graybiel AM,
Swallow JG, Koteja P, Carter PA, Garland T Jr (1999) Artificial
Tonegawa S (1994) Dopamine D1 receptor mutant mice are
selection for increased wheel-running activity in house mice
deficient in striatal expression of dynorphin and in dopamine-
results in decreased body mass at maturity. J Exp Biol
mediated behavioral responses. Cell 79:729–742
B.D.K.C SUPPLEMENTARY REGULATIONS CLUB CHAMPIONSHIPS 2011 Held at Ellough Park Raceway, Beccles, Suffolk, NR34 7XD During 2011 Beccles & District Kart Club Ltd (BDKC, the club) will organize National Clubman Permit meetings on the March, April, May, June, July, August, September, October and November. The meetings will be held under the general regulations of the Motor Sports Assoc
La s ocietà atg tes t s ys tems Gmbh & Co. GG ha s ede in Ger mania, Zum S chlag 3 D-9 7 8 7 7 W er theim. Telefono 0 0 4 9 9 3 4 2 2 9 1 0 ,Fax 0 0 4 9 9 3 4 2 3 9 5 1 0 , s ito W eb. atg-tes t-s ys tems . de. É s tata fondata nel 1 9 7 9 ed ha pr es entato il s uo pr imo s is tema di col audo nel 1 9 8 0 . Oggi è par te del gr uppo Ever et Char les Technologies ed è pr es ente con filial
 and food [Harlan Teklad Laboratory Rodent Diet (8604); aftergeneration 23, breeding females were given Harlan Teklad MouseBreeder Diet (7004)] were available ad libitum. Rooms were con-trolled for temperature (~22°C) and photoperiod 12-h/12-hlight/dark (lights on at 0700 hours, central standard time).
and food [Harlan Teklad Laboratory Rodent Diet (8604); aftergeneration 23, breeding females were given Harlan Teklad MouseBreeder Diet (7004)] were available ad libitum. Rooms were con-trolled for temperature (~22°C) and photoperiod 12-h/12-hlight/dark (lights on at 0700 hours, central standard time).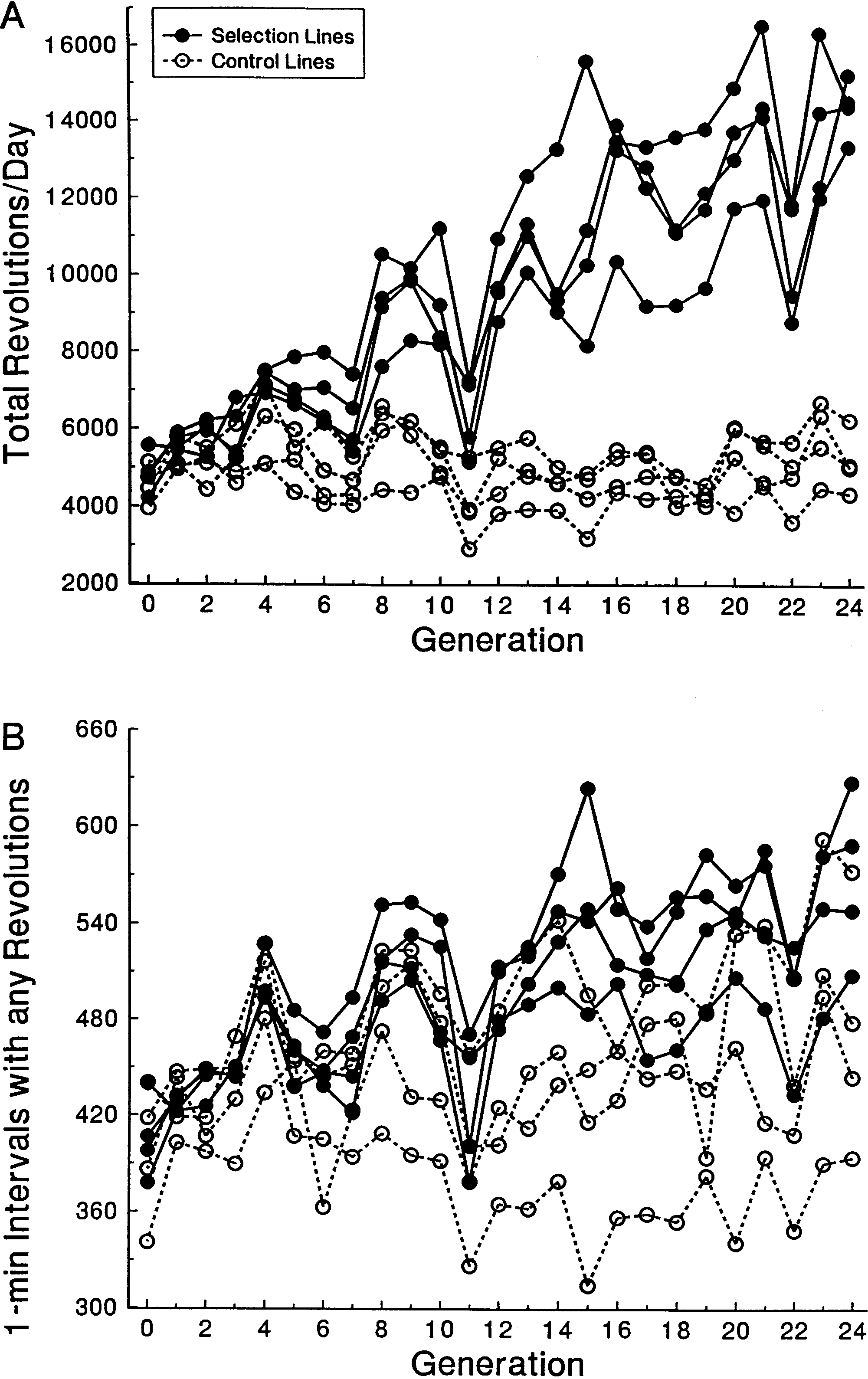 2 days preceding injections for all three drug trials combined. Data were analyzed using a two-way analysis of covariance(ANCOVA), including line type, drug trial, and the interaction be-tween drug trial and line type as cofactors.
2 days preceding injections for all three drug trials combined. Data were analyzed using a two-way analysis of covariance(ANCOVA), including line type, drug trial, and the interaction be-tween drug trial and line type as cofactors.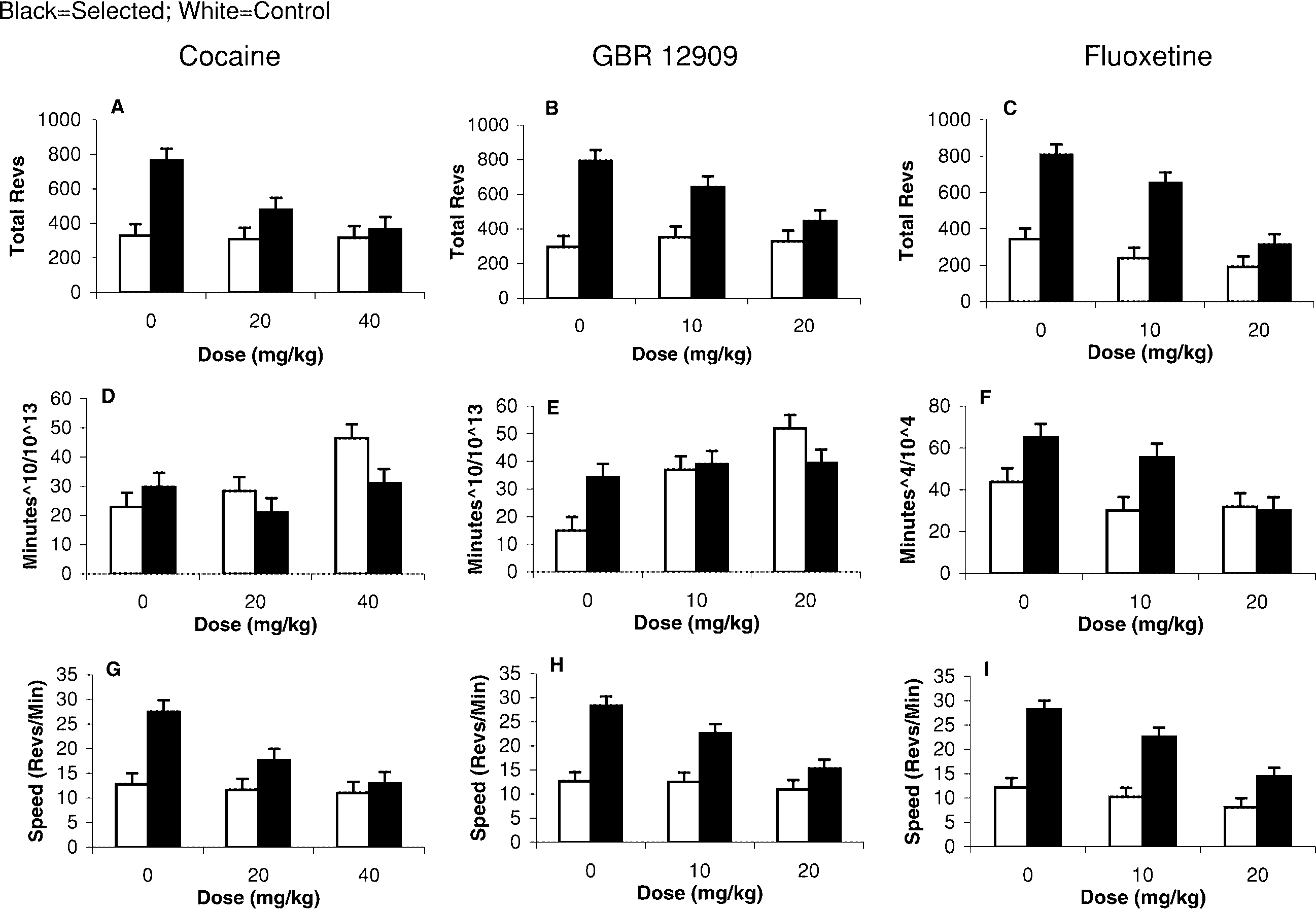 Fig. 3A–I The wheel-running response to i.p. administration of
Fig. 3A–I The wheel-running response to i.p. administration of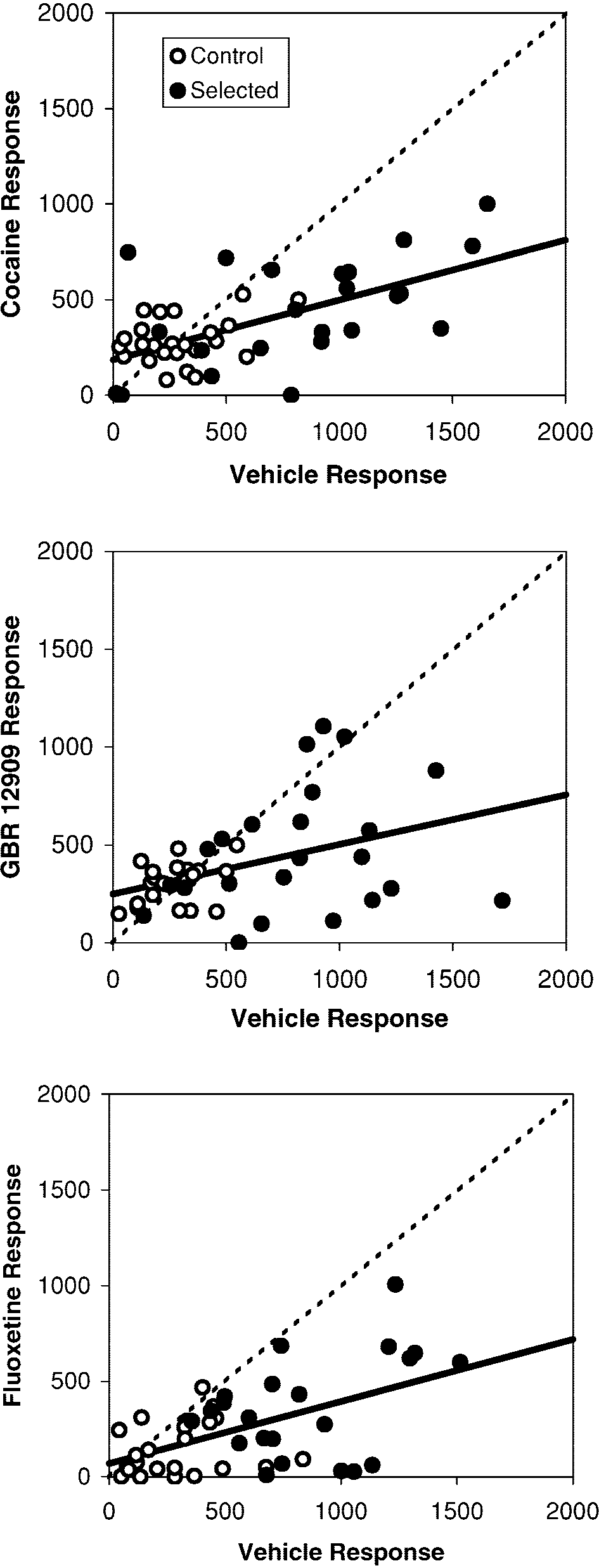 ambulation; P=0.0004; Fig. 5) during the second 24-hperiod of testing. However, there were no significant dif-ferences on the first day of testing (P=0.26 for finemovements and P=0.07 for coarse movements; Fig. 5).
ambulation; P=0.0004; Fig. 5) during the second 24-hperiod of testing. However, there were no significant dif-ferences on the first day of testing (P=0.26 for finemovements and P=0.07 for coarse movements; Fig. 5).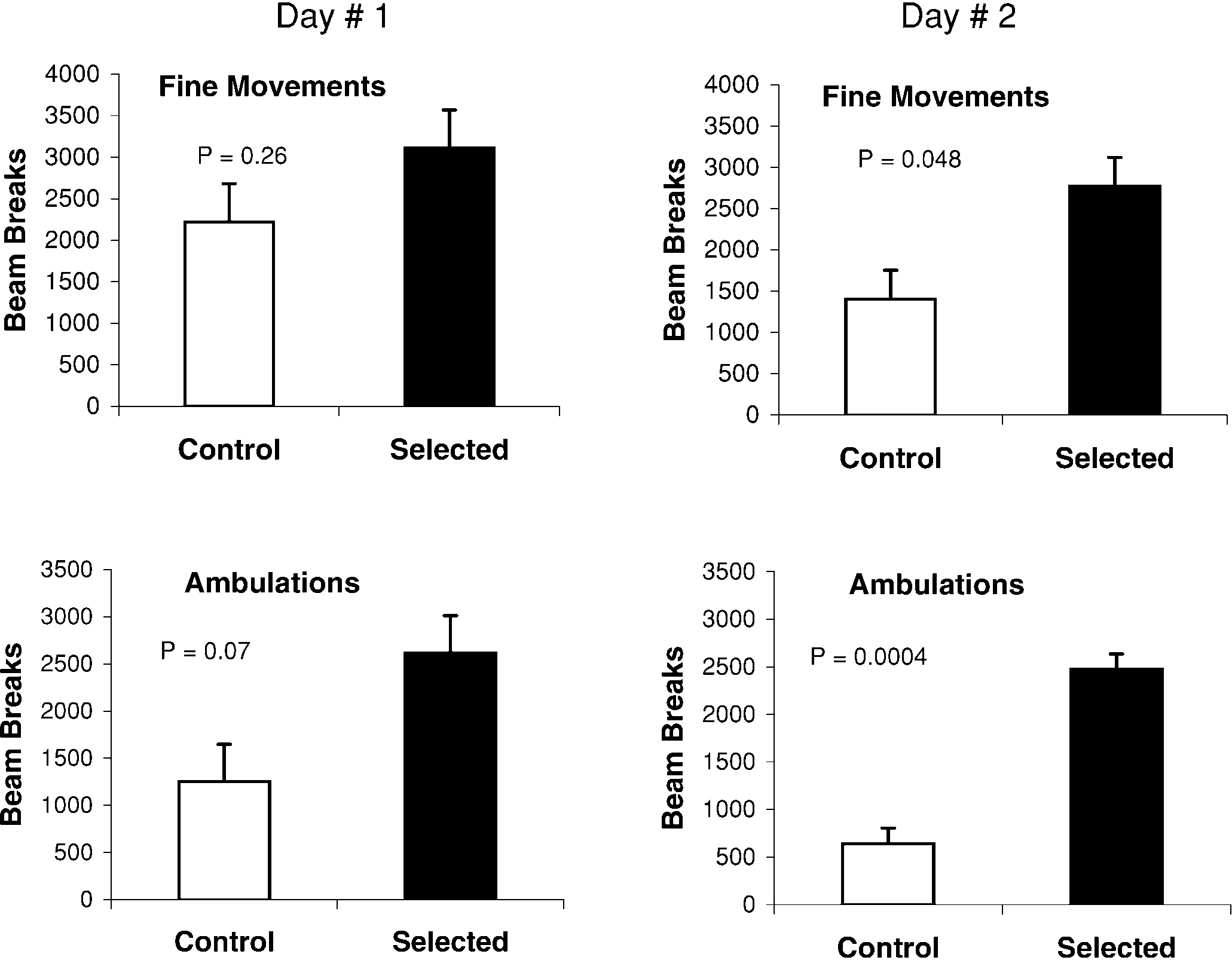 Fig. 5 Activity of mice from
Fig. 5 Activity of mice from