Do you want to buy antibiotics online without prescription? https://buyantibiotics24h.net/ - This is pharmacy online for you!
Pq220012380p
Failure to handle more than one internal representation in visual detection tasks Andrei Gorea*† and Dov Sagi‡
*Laboratoire de Psychologie Expe´rimentale, Centre National de la Recherche Scientifique & Rene´ Descartes University, 71 Ave. Edouard Vaillant, 92774Boulogne-Billancourt, France; and ‡Department of Neurobiology͞Brain Research, Weizmann Institute of Science, Rehovot, 76100 Israel
Edited by Jacob Nachmias, University of Pennsylvania, Philadelphia, PA, and approved August 22, 2000 (received for review April 13, 2000)
Perceptual studies make a clear distinction between sensitivity and
adjusting their decision criteria to the occurrence probability
decision criterion. The former is taken to characterize the process-
of the signal was assessed for a number of different strength
ing efficiency of the underlying sensory system and it increases
signals presented in isolation with one out of three occurrence
with stimulus strength. The latter is regarded as the manifestation
probabilities. A second experimental condition was meant to
of a subjective operation whereby individuals decide on (as op-
check whether the performance above (i.e., both sensitivities
posed to react reflexively to) the occurrence of an event based on
and criteria) is maintained when two equal strength stimuli
factors such as expectation and payoff, in addition to its strength.
with different occurrence probabilities are displayed simulta-
To do so, individuals need to have some knowledge of the internal
neously. Potential context-related interactions between deci-
response distributions evoked by this event or its absence. In a
sion criteria (and sensitivities) were further studied in a third
natural, behaviorally relevant multistimulus environment, observ-
condition involving the simultaneous presentation of stimuli
ers must handle many such independent distributions to optimize
differing in both strength and occurrence probability. their decision criteria. Here we show that they cannot do so. Instead, while leaving sensitivity unchanged, lower and higher Experimental Design and Methods visibility events tend to be reported respectively less and more
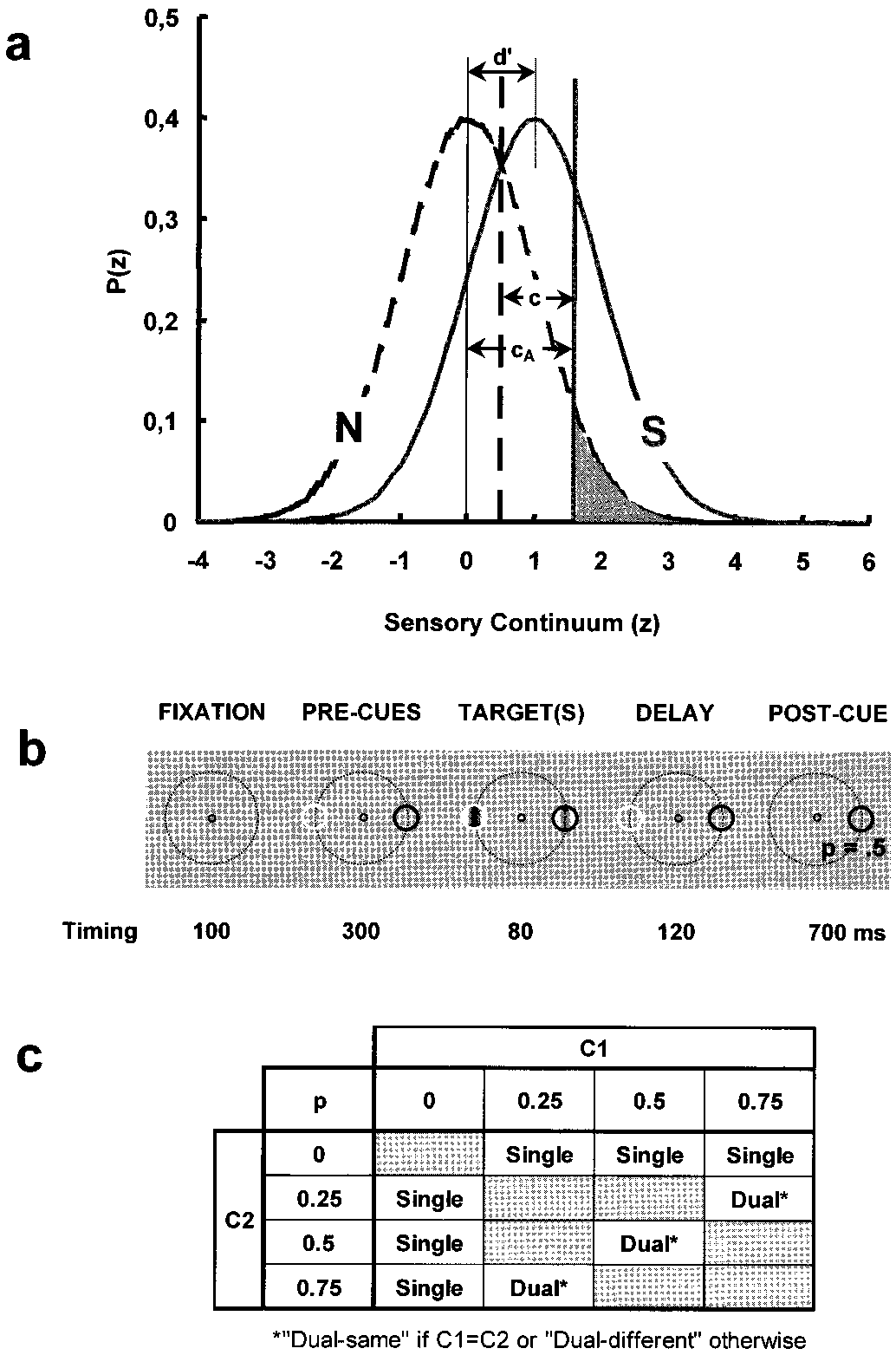
Fig. 1b illustrates the sequence of events within one trial. In each
frequently than when they are presented in isolation. This behav-
block of trials and on each trial, observers were presented with,
ior is in quantitative agreement with predictions based on the
in sequence (i) a fixation white circle present during the whole
notion that observers represent a multistimulus environment as a
trial; (ii) one white and one black ‘‘precue’’ circles; (iii) two 3
unitary internal distribution to which each stimulus contributes
cycles͞deg vertical Gabor targets of contrasts Cproportionally to its probability of occurrence. Perceptual phenom-
sented within the precue circles with probabilities, Pena such as blindsight, hemineglect, and extinction may be, at least
(iv) after their offset (‘‘delay’’ period), one of the precues
in part, accounted for in such a way.
randomly disappeared. Observers had to respond whether or not
a target was presented inside the persistent (postcue) circle only.
decision making ͉ psychophysics ͉ criteria drift ͉ unitary internal
This experimental format combines partial report (4) and signal
detection techniques. It requires that observers monitor both
locations until the ‘‘appearance’’ of the postcue. The stimuli were
It is well established (1, 2) and by now common knowledge presented on a 29 cd͞m2 gray background. The fixation circle
that humans (and nonhumans) can and do set the reference
was 0.5° in diameter. The cue circles were displayed 1.6° to the
strength of an internal response beyond which internal events
left and to the right of fixation (on the dashed large circles absent
will be assumed to represent external events in relation to the
in the actual trials; Fig. 1b) with their locations randomized
probability and͞or the payoff associated with the latter:
equally energetic events will be reported more frequently if
1,P1 and C2,P2 were consistently asso-
ciated with the cue polarities, and observers were informed of
their occurrence is likely (or the payoff is high) than if it is not.
this mapping before each block. A few blocks also were run with
Signal detection experiments also have established that, for a
horizontal targets and͞or with cues and targets displayed above
unique stimulus strength and occurrence probability, observ-
and below fixation. Incorrect responses were signaled by a short
ers can simultaneously entertain a number of response criteria
that translate into different levels of confidence as to the
For each experimental block, the contrasts of the two targets
occurrence of that stimulus (1, 2). The literature remains
silent, however, on the issue of whether or not humans can
1 and C2) and their probabilities (P1 and P2) were fixed and
systematically mapped on each of the two cue polarities with
entertain multiple criteria based on different probabilities
observers having full knowledge of this cue-color͞stimulus-
associated with a number of simultaneous, spatially distinct
mapping at the start of each experimental block. C
events. Common sense admits the proposition according to
in the range of 0.02 to 0.05. The specific values for each observer
which the (yes͞no) report of the occurrence of any among N
were chosen based on preliminary experiments with no other
distinct events can be biased by the a priori likelihood of that
event (Fig. 1a). But is this still the case in an environment
Јs between 0.5 and 3.5 (i.e., most of the
characterized by a variety of stimulus intensities? A multi-
Ј range) and dЈ differences between 0.7 and 2 (see
stimulus environment where different events differ in their
below). Probabilities of the targets could be 0, 0.25, 0.50, or 0.75.
likelihood is the typical milieu of most organisms. Character-
Stimulus characteristics were manipulated to yield three exper-
izing decision making within such an ambience is, then, of
critical significance for understanding how humans regulate
The ‘‘single’’ condition involved the presentation of one single
(or adapt) the point on their sensory continuum they use as a
target in each trial (obtained by setting one of the two proba-
reference for judging ‘‘reality’’ given the actual magnitudes
and probabilities of the ongoing physical events (3). In the
This paper was submitted directly (Track II) to the PNAS office.
present study, this broad issue is scaled down to its most
†To whom reprint requests should be addressed. E-mail: gorea@psycho.univ-paris5.fr.
elementary form, whereby decision criteria are studied for
only two stimuli at a time that may or may not differ in their
The publication costs of this article were defrayed in part by page charge payment. Thisarticle must therefore be hereby marked “advertisement” in accordance with 18 U.S.C.
strength. In one ‘‘baseline’’ condition, observers’ capacity of
§1734 solely to indicate this fact. 12380 –12384 ͉ PNAS ͉ October 24, 2000 ͉ vol. 97 ͉ no. 22
Under the ‘‘dual’’ conditions, two stimuli͞signals were pre-
sented in each trial with their probabilities paired so as to yield
a constant average signal probability of 0.5 (i.e., 0.25–0.75,
0.50–0.50, and 0.75–0.25). Thus, here again three dЈ and cA
values were assessed for each contrast in a pair. There were two
sorts of dual conditions. Under the ‘‘dual-same’’ condition, the
two stimuli in a pair were of identical contrasts (C1 ϭ C2). Under
the ‘‘dual-different’’ condition, the two contrasts were different
(C1 Ͻ C2) and paired so as to yield dЈ differences between 0.7 and
2. Fig. 1c displays the 12 stimulus pairings defined by the
probabilities and contrasts of the stimuli. C-P pairings were
The most extensive data sets (three contrast pairs) were
obtained from two naive, well-trained observers. An additional
naive observer VR and the first author were run with only one
contrast pair. Some critical points in the main experiment also
were confirmed with the second author. Observer AG and
additional naive observer VR also were run in a second exper-
iment equivalent in all respects to the dual-different condition
with the exception that one of the two stimuli never had to be
reported and the observers were told so from the start. In all
cases, one datum point was estimated from at least 312 trials but
more typically from 416–832 trials. Results Operationally, the question raised here is whether or not the
criteria for the paired stimuli remain unchanged when assessed
under single and dual conditions. Fig. 2a shows that the absolute
PSYCHOLOGY
criteria (cA ϭ ϪzFA, i.e., the standard z-score of false alarms;
Fig. 1a) for single and dual-same conditions are scattered about
the identity (slope 1, dotted) line for both the low (C1, solid
symbols; paired t test ϭ 0.3, df ϭ 40, P ϭ 0.38, not significant)
and the high (C2, open symbols; paired t test ϭ Ϫ1.05, df ϭ 40,
P ϭ 0.15, not significant) contrasts used with each observer. The
linear regression lines with errors in both coordinates (5) fit to
each data subset (solid and dashed lines, respectively) are also
close to this identity line. For both single and dual-same con-
ditions, the average likelihood ratio criterion (; Fig. 1a) for
signal probabilities 0.25, 0.5, and 0.75 was (in agreement with the
(a) The Signal Detection Theory (1) framework for the dual-criterion
known conservative behavior of humans; ref. 1) 1.35, 1.05, and
experiment. Gaussian functions describe the probability density, P(z), of theinternal response distributions (in standard z-scores; abscissa) for the noise [N:
0.75 and did not depend on dЈ. This is evidence that, for spatially
PN(z)] alone (dashed curve) and for the signal ϩ noise [S: PS(z)]. Thin vertical
distinct, equal strength stimuli, observers can and do entertain
lines show their means with sensitivity (dЈ ϭ zHit Ϫ zFA, with zHit and zFA the
independent criteria in relation to the specified stimulus
z scores for the observed correct target detection and false alarm rates) being
the distance between these means (dЈ ϭ 1 in this case) measured in units of the
In contrast with the above, cAs assessed under the dual-
noise standard deviation, N, and assuming that N and S are normally distrib-
different condition are systematically different from the corre-
uted with ϭ N ϭ S. We define an ‘‘absolute’’ criterion as cA ϭ ϪzFA.
sponding ones observed under the single and dual-same condi-
Defined in this way, criteria are independent of the univariance assumption
tions (Fig. 2b). With very few exceptions, they lie above (solid
(i.e., S ϭ N), because they depend on the N distribution only. The corre-
sponding values of the likelihood ratio criterion,
ϭ Ϫ5.03, df ϭ 46, P Ͻ 0.00002) and below
 ϭ Ps(z ϭ cA)͞PN(z ϭ cA),
characterize observers’ response bias independently of dЈ. Error rate is mini-
(open symbols; paired t test ϭ 4.37, df ϭ 46, P Ͻ 0.0001) the
mized when  ϭ PN͞PS, (with PN and PS the a priori N and S probabilities) but
identity (slope 1, dotted) line for the low (C1) and high (C2)
experimental results show that observers adopt a more conservative behavior
contrasts, respectively. This is to say that the cAs associated with
with s closer to one (1). The vertical dashed and continuous heavy lines show
the low and high contrasts in a dual-different pair ‘‘drift’’ as if
optimal criteria for PS ϭ 0.5 and PS ϭ 0.25, respectively. The shaded area
‘‘attracted’’ by the mean of the internal response distribution
denotes the False Alarm (FA) rate for the latter case. (b) One trial sequence as
associated with the high and low contrast stimuli, respectively.
detailed in the text. (c) The 12 experimental conditions as characterized by the
This is so despite the fact that observers have full knowledge of
combination of two distinct stimuli of contrast C1 and C2 and of four stimulus
the properties of the signals, that is, of their contrasts and
Fig. 2 c and d shows that, unlike cAs, dЈs remain insensitive to
bilities, or equivalently, one of the two contrasts in a pair to zero).
the experimental condition. They lie close to the slope 1
Stimulus characteristics (C and P) were fixed and announced at
diagonal, whether assessed in single vs. the average of the
the beginning of each experimental block. The target was
dual-same and dual-different conditions (open and solid sym-
presented randomly across trials within a white or a black cue
bols, respectively, in Fig. 2c) or under dual-same vs. dual-
circle so that cue polarity yielded no meaning in this condition.
different conditions (Fig. 2d).
For a given C, the probability of the signal (0.25, 0.50, or 0.75)
The cA shifts observed under the dual-different condition are
was randomized across blocks so that three sensitivity (dЈ) and
not entailed by the mere presence of two targets of different
criterion (cA, ; Fig. 1a) values were assessed for each C. Signal
contrasts, but rather by the decisions taken on their occurrence.
contrasts also were randomized across blocks.
The dual-different condition was rerun with two observers (AG
PNAS ͉ October 24, 2000 ͉ vol. 97 ͉ no. 22 ͉ 12381
Correspondence between criteria (a and b) and dЈs (c and d) assessed under single, dual-same, and dual-different conditions. Different symbols are for
different observers. Data points represented by identical symbols are for different stimulus probabilities and͞or contrasts. (a and b) Solid and open symbols showthe absolute criteria obtained for the low (C1) and high (C2) contrasts, respectively, in a stimulus pair. (a) cA[single] vs. cA[dual-same] comparison. The identitylocus is shown as the dotted (slope 1) line. Solid and dashed regression lines are fits with errors in both coordinates (5) through the solid (C1 stimuli) and open(C2 stimuli) symbols, respectively. (b) As in a but for the cA[mean (single ϩ dual-same)] vs. cA[dual-different] comparison. Note the upward and downward shiftsof the dual-different criteria associated with low (solid symbols) and high (open symbols) contrasts, respectively, in a stimulus pair. (c) dЈ[single] vs. dЈ[dual]comparison with closed (solid regression line) and open (dashed regression line) symbols showing dual-different and dual-same data, respectively. The identityline is dotted. (d) dЈ[dual-same] vs. dЈ[dual-different] comparison. The identity and the regression line through all experimental points are dotted and solid,respectively.
and VR) under conditions where one of the two stimuli (the
representation of the internal response distributions associated
‘‘companion’’) was never reported. The postcue was never
with each external event is of critical importance for the effi-
presented for this stimulus, whereas the two cue polarities were
ciency of these decision processes. Decision criteria should be set
randomized over the two stimulus contrasts. For a target stim-
so as to minimize some cost function, for example, the number
ulus of 4%, observer VR showed a slight but consistent criterion
of response errors (1). A failure to correctly represent the
drop when the probability of either a 6% or 60% contrast
distinct distributions associated with the different targets would
companion (used in different blocks of trials) increased from
translate into criteria shifts. The presently observed criteria
zero to one. This is an effect opposite to the one observed in the
drifts may be regarded as the consequence of such a failure.
main experiment. Observer AG showed no interaction whatso-
More specifically, the present data are well fit by a model with
ever for either a 4% or 6% target in the presence of either a 5%
no free parameters posing that observers have access to only one
or 60% companion (with each contrast pair run in different
internal response distribution generated by the incoming stimuli.
blocks). Thus, when one internal response distribution needs not
This unitary distribution is the weighted sum of the internal
to be estimated (because no related report is requested), the
distributions evoked by each of these stimuli with weights equal
criteria used for the target stimuli do not drift toward the mean
to their respective occurrence probabilities. Observers decide on
of that distribution (as in the main experiment). This observation
the occurrence of a signal in the standard way, that is, by
goes against an interpretation of the main results reported here
reference to a given likelihood ratio criterion (), a number that
in terms of some sort of sensory adaptation (6).
depends only on the signal probability as denoted by the
associated precues, and for that matter, postcues. However, the
Modeling the Data and Discussion
likelihood ratios observers use for the comparison with this
In a multistimulus environment where, potentially, decisions are
number are computed from the merged distribution. For equal
to be made on the occurrence of any external event, the reliable
strength stimuli, the unitary distribution will be identical to each
12382 ͉ www.pnas.org PSYCHOLOGY
Correspondence between data and predictions by a dual and unitary internal distribution model. (a) cA differences (⌬cA) measured in the dual-different
condition (abscissa) are compared with the corresponding ⌬cAs averaged over the single and dual-same conditions (open symbols and dashed line; two-distribution model) and with the predictions of the unitary distribution model (closed symbols). ⌬cAs were computed as cA(C2,P) Ϫ cA(C1,1 Ϫ P), that is betweenthe criteria associated with the high contrast (C2) signal of probability P and the low contrast (C1) signal of probability 1 Ϫ P. Circles: cA(C2,0.25) Ϫ cA(C1,0.75);squares: cA(C2,0.50) Ϫ cA(C1,0.50); triangles cA(C2,0.75) Ϫ cA(C1,0.25); note that in the dual-different condition, the above predicted (unitary distribution) andmeasured ⌬cAs are, in order, small and negative, close to zero, and large and positive. Symbols lying along the diagonal of slope 1 passing through the originshow perfect correspondence between data and predictions. The main assumption of the unitary model (b–d) is that, in a multistimulus environment, observersset, for a given stimulus probability, the same (nonoptimal) likelihood ratio criterion () as in a single-stimulus (or uniform) environment whereas the likelihoodratios they use for the comparison with this number are computed from the unitary internal response distribution (heavy continuous curves) generated by theensemble of stimuli on which they have to decide. The unitary distribution is the weighted sum of the internal distributions evoked by each of these stimuli withweights equal to their respective occurrence probabilities (thin Gaussian curves). (b–d) The three contrast-probability pairings used in the experiments. Theweaker and stronger signals in a pair are presented with probabilities 0.25, 0.75 (a), 0.5, 0.5 (b), and 0.75, 0.25 (c). Heavy vertical lines show where the criteriashould be for each of these probabilities assuming that observers are optimal and use the same  as in the single and͞or dual-same conditions. In practice,observers display a conservative behavior under all conditions so that these criteria are closer to each other than shown. Vertical dotted lines show the meansof the N- and the two S-distributions. The actual internal response distributions (N and S) remain unchanged under all experimental conditions, so that a giventarget yields a constant dЈ across all conditions.
individual distribution so that the predicted response criteria for
criteria as those assessed under the single and͞or dual-same
such dual-same conditions should not differ from those observed
conditions. The open symbols in Fig. 3a represent the actual
under single conditions. Note that this model bears only on how
⌬cAs between the same stimuli as measured in the single and͞or
observers use the internal response distributions in the decision
dual-same cases. Overall, ⌬cAs under the dual-different condi-
process whereas leaving these distributions, and therefore, sen-
tion are smaller than those under the single and dual-same
sitivity, unaffected by the experimental manipulations. Indeed,
conditions (‘‘attraction’’; Fig. 2b) by an average of 0.44 (for an
the present data show no systematic dЈ changes with these
In a multistimulus environment, precues (and for that matter,
In Fig. 3a, the ‘‘unitary distribution’’ model is used to predict
postcues) are helpful for optimizing performance when they are
the cA differences (⌬cA) between the two stimuli in the dual-
used to tag stimulus probabilities but not stimulus strengths.
different pairs (closed symbols). This was achieved by using the
Observers decide on the presence͞absence of a signal in relation
observers’ dЈs and s as assessed for the same stimuli and
to the internal response distribution associated with a given
stimulus probabilities as under the single and dual-same condi-
external event (7). Optimal decisions in the presence of more
tions. Fig. 3 b and c provides an illustration of how this was
than one stimulus require the appraisal of the internal events
accomplished (see legend). Predictions of the unitary distribu-
distributions associated with each stimulus. The present data
tion model lie very close to the line of perfect fit (the diagonal
show that human observers cannot do so even though the
of slope 1) and account for 80% of the variance. On the
different signals are spatially disjoint and unambiguously de-
alternative, standard Signal Detection Theory assumption that
noted by consistent precues. Instead, observers appear to use the
observers keep track of and independently use each of the two
composite distribution generated by the two simultaneously
internal distributions associated with the two stimuli under the
occurring stimuli and use it as a unitary distribution to set their
dual conditions, the dual-different case should yield the same
criteria in accordance with the specified signal probabilities as
PNAS ͉ October 24, 2000 ͉ vol. 97 ͉ no. 22 ͉ 12383
(in)efficiently as in a single-stimulus environment. The global
different strength signals are randomly mixed in one experimen-
effect of this behavior is that, relative to the impoverished
tal block. Not knowing the specific signal (among many) to which
(single-strength) environments, the criteria they use in more
an internal response distribution should be associated, observers
ecological ones are displaced to higher values on the sensory
have no means of keeping track of these distributions; as a
scale (more misses) for the weaker signals, and to lower values
consequence, they place their detection or classification criteria
(more false alarms) for the stronger ones, provided that the
in relation to a global internal response distribution representing
external events considered are behaviorally relevant. This fact
the ensemble of relevant stimuli. In the present experimental
generalizes to conditions where the paired stimuli are of orthog-
format, however, observers could and actually did keep perfect
onal orientations or different spatial frequencies, or are pre-
track of the stimulus identity by using the polarity of the precue
and postcue circles. This is indicated by the fact that the criteria
The present results may bear on the well-known neuropsy-
obtained under the single and dual-same conditions for the same
chological disorders of blind-sight (9), hemineglect (10), and
signal probabilities are practically identical (Fig. 2a). Why then
extinction (11). Consequences of cortical injury, these conditions
couldn’t observers adjust their criteria in relation to each of the
refer, respectively, to the relatively efficient behavior toward
two distinct internal response distributions generated under the
objects which the patients claim not to see, to their neglect of half
of their sensory space, or of some simultaneously occurring
One may pose that, whatever the sampling resources of the
stimuli. Presumably, patients of this kind are in a situation
system as a whole, it devotes them all to sampling one (intensive)
analogous to the one studied here. Given that they present a
reality at a time to reach a decision bearing on this reality. If
critical sensitivity drop contralateral to the injury, and on the
equivalent criterion shifts were to be observed within the context
assumption of a homogeneous physical world, such patients
of discrimination tasks along dimensions other than intensity
might displace their response criterion toward the mean of their
(e.g., shape, color, and velocity), this could be regarded as a
internal responses along the nonaffected sensory space. They
fundamental constraint imposed by the unity of action (or
will then ignore the weaker internal responses generated in the
decision) on this reality (17). An alternative interpretation of the
affected areas. Criterion shifts under these neurological condi-
present results would be that the sampling resources of the brain
tions have been invoked (12, 13) and actually observed (14).
are limited [e.g., to 7 Ϯ 2 samples or chunks (18) or less (8)] and
The criterion shifts assessed in the present study depend on
that they cannot be distributed across external events without a
both the specific strengths (or dЈs) and the probabilities of the
significant loss in accuracy. One way or another, the global use
stimuli involved in the detection task. They are quantitatively
of a unitary internal response distribution in judging multiple
accounted for by a model whereby, for a given stimulus proba-
simultaneously occurring events might account for the unity of
bility, observers use the same likelihood ratio criterion in a
the (visual) world as experienced and as acted on. Of course,
single-stimulus (uniform) or multistimulus environment but
the generality of this statement is subject to further studies
compute the current likelihood ratios needed for the comparison
of the present phenomenon in a variety of multidimensional
with this number from the unitary internal response distribution
generated by the ensemble of stimuli on which they have to
decide. This behavior is to be expected in detection (15) or
We thank Randolph Blake, Patrick Cavanagh, Elliot Freeman, Avi
scaling͞classification experiments (6, 16) where nontagged,
Karni, Stanley Klein, Larry Maloney, and Joshua Solomon for valuable
comments on earlier versions of this paper and Yoram Bonneh for
providing the software. This work was supported in part by a joint
§Gorea, A. & Sagi, D. (2000) Investig. Ophtal. Vis. Sci. Suppl., 41, S751 (abstr.).
French-Israeli Grant ‘‘Arc-en-ciel.’’
1. Green, D. M. & Swets, J. A. (1974) Signal Detection Theory and Psychophysics
10. Robertson, I. H. & Marshall, J. C. (1994) Unilateral Neglect: Clinical andExperimental Studies (Brain Damage and Cognition) (Robin Walker, London).
2. Macmillan, N. A. & Creelman, C. D. (1991) Detection Theory: A User’s Guide
11. Humphreys, G. W., Romani, C., Olson, A., Riddoch, M. J. & Duncan, J. (1988)
(Cambridge Univ. Press, Cambridge, U.K.). Nature (London) 372, 357–359.
3. Tversky, A. & Kahaneman, D. (1974) Science 185, 1124–1131.
12. Campion, J., Latto, R. & Smith, Y. M. (1983) Behav. Brain Sci. 3, 423–448.
4. Sperling, G. (1960) Psychol. Mono. 74, 1–29.
13. Klein, S. A. (1998) in Toward a Science of Consciousness II: The Second Tucson
5. Press, W. H., Teukolsky, S. A., Vetterling, W. T. & Flannery, B. P. (1992)
Discussions and Debates, eds. Hemeroff, S. R., Kaszniak, A. W. & Scott, A. C. Numerical Recipes in C (Cambridge Univ. Press, Cambridge, U.K.), pp.
(MIT Press, Cambridge MA), pp. 361–369.
14. Azzopardi, P. & Cowey, A. (1998) Consciousness Cognition 7, 292–311.
15. Gescheider, G. A., Wright, J. H., Weber, B. J., Kirchner, B. M. & Milligan, E. A.
6. Helson, H. (1964) Adaptation-Level Theory (Harper & Row, New York).
(1969) Percept. Psychophys. 5, 18–20.
7. Vickers, D. (1979) Decision Processes in Visual Perception (Academic, New
16. Parducci, A. (1974) in Handbook of Perception: Psychophysical Judgement andMeasurement, eds. Carterette, E. C. & Friedman, M. P. (Academic, New York),
8. Cowan, N. (2001) Behav. Brain Sci., in press.
9. Weiskrantz, L. (1997) Consciousness Lost and Found: A Neuropsychological
17. Cotterill, R. M. J. (1995) J. Consciousness Studies 2, 290–312. Exploration (Oxford Univ. Press, Oxford).
18. Miller, G. A. (1956) Psychol. Rev. 63, 81–97. 12384 ͉ www.pnas.org
Please read the following instructions carefully The examination is of two hours’ duration. Candidates are required to answer FOUR questions, TWO from PART A and TWO from PART B . All four questions are of equal value. Answers must be fully reasoned and supported by authority where appropriate. Candidates need to take care to read the questions carefully and to answer what is
CAMAG APPLICATION NOTES ARRANGED IN CLASSIFICATION SECTIONS I Quantitative determinations Biochemical research / biotechnology A-51.1 - Methionine (in fermentation broth) A-76.1 - Rape seed oil in fermentation broth Clinical A-01.4 - Carbamazepine and two of its metabolites in serum A-04.2 - Diazepam trace analysis of by-products in the ppm range * A-21.1 - Inorganic and or
 Under the ‘‘dual’’ conditions, two stimuli͞signals were pre-
sented in each trial with their probabilities paired so as to yield
a constant average signal probability of 0.5 (i.e., 0.25–0.75,
0.50–0.50, and 0.75–0.25). Thus, here again three dЈ and cA
values were assessed for each contrast in a pair. There were two
sorts of dual conditions. Under the ‘‘dual-same’’ condition, the
two stimuli in a pair were of identical contrasts (C1 ϭ C2). Under
the ‘‘dual-different’’ condition, the two contrasts were different
(C1 Ͻ C2) and paired so as to yield dЈ differences between 0.7 and
2. Fig. 1c displays the 12 stimulus pairings defined by the
probabilities and contrasts of the stimuli. C-P pairings were
The most extensive data sets (three contrast pairs) were
obtained from two naive, well-trained observers. An additional
naive observer VR and the first author were run with only one
contrast pair. Some critical points in the main experiment also
were confirmed with the second author. Observer AG and
additional naive observer VR also were run in a second exper-
iment equivalent in all respects to the dual-different condition
with the exception that one of the two stimuli never had to be
reported and the observers were told so from the start. In all
cases, one datum point was estimated from at least 312 trials but
more typically from 416–832 trials.
Under the ‘‘dual’’ conditions, two stimuli͞signals were pre-
sented in each trial with their probabilities paired so as to yield
a constant average signal probability of 0.5 (i.e., 0.25–0.75,
0.50–0.50, and 0.75–0.25). Thus, here again three dЈ and cA
values were assessed for each contrast in a pair. There were two
sorts of dual conditions. Under the ‘‘dual-same’’ condition, the
two stimuli in a pair were of identical contrasts (C1 ϭ C2). Under
the ‘‘dual-different’’ condition, the two contrasts were different
(C1 Ͻ C2) and paired so as to yield dЈ differences between 0.7 and
2. Fig. 1c displays the 12 stimulus pairings defined by the
probabilities and contrasts of the stimuli. C-P pairings were
The most extensive data sets (three contrast pairs) were
obtained from two naive, well-trained observers. An additional
naive observer VR and the first author were run with only one
contrast pair. Some critical points in the main experiment also
were confirmed with the second author. Observer AG and
additional naive observer VR also were run in a second exper-
iment equivalent in all respects to the dual-different condition
with the exception that one of the two stimuli never had to be
reported and the observers were told so from the start. In all
cases, one datum point was estimated from at least 312 trials but
more typically from 416–832 trials.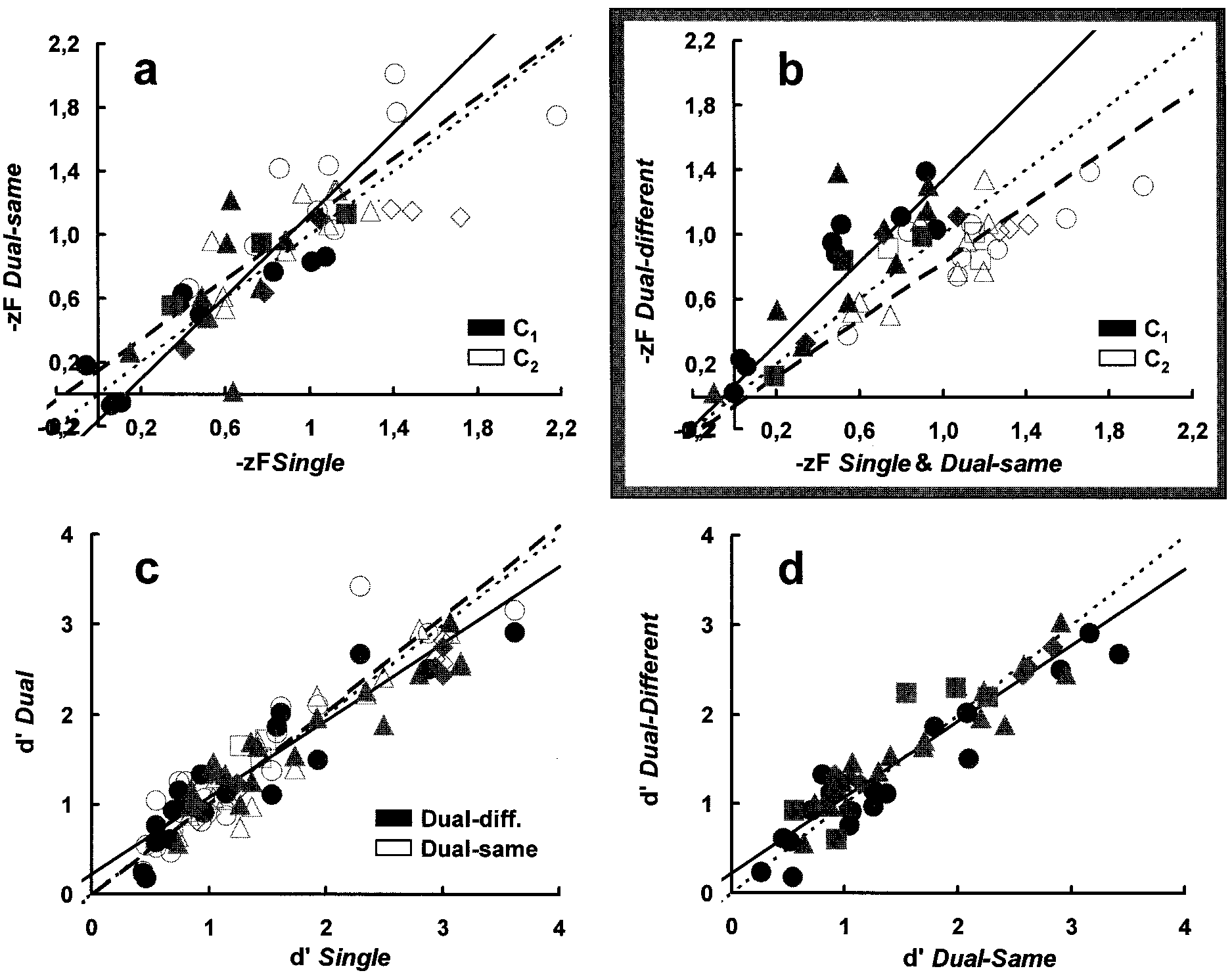 Correspondence between criteria (a and b) and dЈs (c and d) assessed under single, dual-same, and dual-different conditions. Different symbols are for
different observers. Data points represented by identical symbols are for different stimulus probabilities and͞or contrasts. (a and b) Solid and open symbols showthe absolute criteria obtained for the low (C1) and high (C2) contrasts, respectively, in a stimulus pair. (a) cA[single] vs. cA[dual-same] comparison. The identitylocus is shown as the dotted (slope 1) line. Solid and dashed regression lines are fits with errors in both coordinates (5) through the solid (C1 stimuli) and open(C2 stimuli) symbols, respectively. (b) As in a but for the cA[mean (single ϩ dual-same)] vs. cA[dual-different] comparison. Note the upward and downward shiftsof the dual-different criteria associated with low (solid symbols) and high (open symbols) contrasts, respectively, in a stimulus pair. (c) dЈ[single] vs. dЈ[dual]comparison with closed (solid regression line) and open (dashed regression line) symbols showing dual-different and dual-same data, respectively. The identityline is dotted. (d) dЈ[dual-same] vs. dЈ[dual-different] comparison. The identity and the regression line through all experimental points are dotted and solid,respectively.
Correspondence between criteria (a and b) and dЈs (c and d) assessed under single, dual-same, and dual-different conditions. Different symbols are for
different observers. Data points represented by identical symbols are for different stimulus probabilities and͞or contrasts. (a and b) Solid and open symbols showthe absolute criteria obtained for the low (C1) and high (C2) contrasts, respectively, in a stimulus pair. (a) cA[single] vs. cA[dual-same] comparison. The identitylocus is shown as the dotted (slope 1) line. Solid and dashed regression lines are fits with errors in both coordinates (5) through the solid (C1 stimuli) and open(C2 stimuli) symbols, respectively. (b) As in a but for the cA[mean (single ϩ dual-same)] vs. cA[dual-different] comparison. Note the upward and downward shiftsof the dual-different criteria associated with low (solid symbols) and high (open symbols) contrasts, respectively, in a stimulus pair. (c) dЈ[single] vs. dЈ[dual]comparison with closed (solid regression line) and open (dashed regression line) symbols showing dual-different and dual-same data, respectively. The identityline is dotted. (d) dЈ[dual-same] vs. dЈ[dual-different] comparison. The identity and the regression line through all experimental points are dotted and solid,respectively.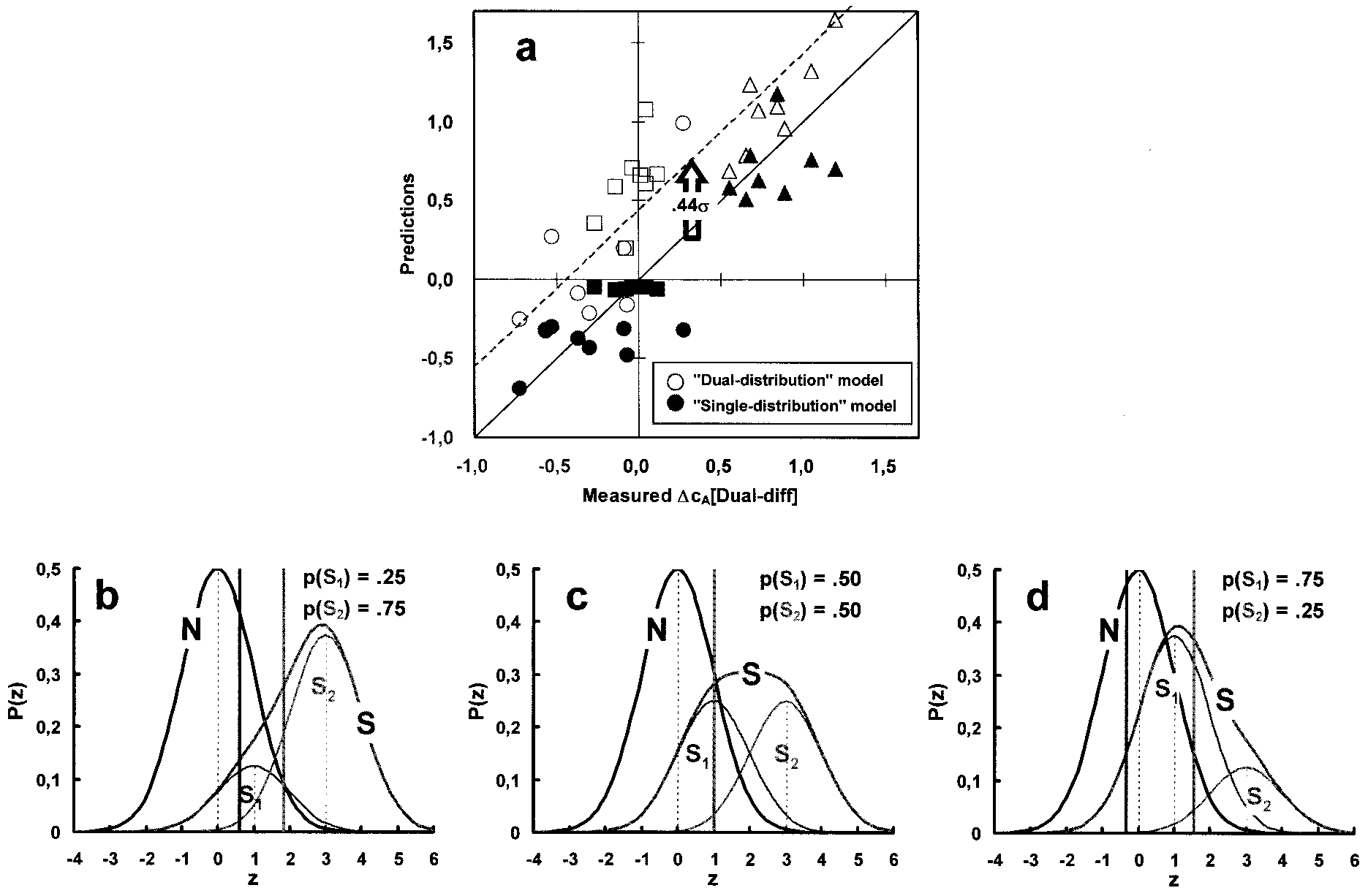 PSYCHOLOGY
PSYCHOLOGY